Навигация
3. Phagemids
Phagemids combine desirable properties of both plasmids and filamentous phages. They carry
· the ColEl origin of replication,
· a selectable marker such as antibiotic resistance,
· the major intergenic region of a filamentous phage .
The segments of foreign DNA cloned in these vectors can be propagated as plasmids. When cells harboring these plasmids are infected with a suitable helper bacteriophage, the mode of replication of the plasmid changes under the influence of the gene II product of the incoming virus.
Interaction of the intergenic region of the plasmid with the gene II protein initiates the rolling-circle replication to generate copies of one strand of the plasmid DNA, which are packaged into progeny bacteriophage particles. The single-stranded DNA purified from these particles is used as a template to determine the nucleotide sequence of one strand of the foreign DNA segment, for site-directed mutagenesis or as a strand-specific probe. Phagemids provide high yields of double-stranded DNA and render unnecessary the time-consuming process of subcloning DNA fragments from plasmids to filamentous bacteriophages.
4. Bacteriophage Vectors
Both single-stranded (filamentous) and double-stranded E.coli phages have been exploited as cloning vectors.
Frederick Twort (1915) and Felix d’Herelle (1917) were the first to recognize viruses which infect bacteria, which d'Herelle called bacteriophages (eaters of bacteria). [7]
Figure 5. Frederick Twort and Felix d’Herelle


4.1 Filamentous phages
Filamentous phages are not lytic. They coexist with the infected cells for several generations and are convenient for cloning genes which produce toxic products. Among the filamentous phages, fd, fl, and M13 have been well characterized and their genomes have been sequenced [4]. Their gene functions and molecular mode of propagation are very similar. They infect cells via F pili, and the first mature phage appears within 15 min [6].
Phage M13 is widely used in nucleotide sequencing and site-directed mutagenesis since its genome can exist either in a single-stranded form inside a phage coat or as a doublestranded replicative form within the infected cell. During replication, only the plus strand of the replicative form is selectively packaged by the phage proteins [1]. The replicative form is a covalently closed circular molecule and hence can be used as a plasmid vector and transformed into the host by the usual transformation procedures. The vectors derived from M13, have the same polylinker as that of pUC18 and pUC19, respectively [2]. The DNA fragments having noncomplementary ends can be directionally cloned in this pair of vectors, and the two strands of DNA can be sequenced independently.
4.2.Double-stranded phage vectors
Of the double-stranded phages, bacteriophage lambda-derived vectors are the most popular tools for several reasons:
· acceptance by the phage of large foreign DNA fragments, thereby increasing the chances of screening a single clone carrying a DNA sequence corresponding to a complete gene;
· development and availability of refined techniques aimed at minimizing the problems of background due to nonrecombinants;
· the possibility of screening several thousand clones at a time from a single petri plate; and, finally,
· the ease with which the phage library can be stored as a clear lysate at 4°C for months without significant loss in plaque-forming activity [7].
Recently, a bacteriophage P1 cloning system has been developed which permits cloning of DNA fragments as large as 100 kbp with an efficiency that is intermediate between cosmids and yeast artificial chromosomes .
5. Scope of Present Review
The extensive knowledge of the basic biology of lambda has permitted modifications of its genome to suit the given experimental conditions. In the present review we describe how the utility of lambda as a cloning vector rests essentially in its intrinsic molecular organization. The following sections give an account of various problems encountered in constructing lambda vectors and the strategies that have been adopted to overcome them. A few commonly used vectors are described in detail, taking into account their special values and limitations. The different methods for screening and storage of genomic and cDNA libraries in lambda vectors are also discussed.
6. Life cycle and genetics of Lambda
An understanding of the basic biology of lambda, its mode of propagation, and the genetic and molecular mechanisms that control its life cycle is needed before its applications for genetic manipulations are discussed. This section deals with the basic biology of lambda.
The lambda virus particle contains a linear DNA of 48,502 bp with a single-stranded 5' extension of 12 bases at both ends; these extensions are complementary to each other.
These ends are called cohesive ends or cos. During infection, the right 5' extension (cosR), followed by the entire genome, enters the host cell. Both the cos ends are ligated by E. coli DNA ligase, forming a covalently closed circular DNA which is acted upon by the host DNA gyrase, resulting in a supercoiled structure.
Похожие работы
... enable the seeds to survive the winter. Overwintering might allow the plant to become a weed or might intensify weedy properties it already possesses. Change in Herbicide Use Patterns Crops genetically engineered to be resistant to chemical herbicides are tightly linked to the use of particular chemical pesticides. Adoption of these crops could therefore lead to changes in the mix of chemical ...
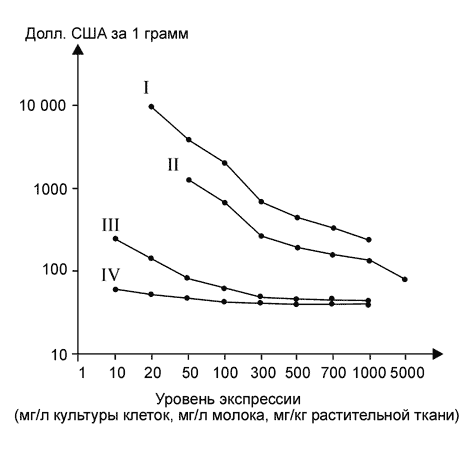
... растений позволит значительно снизить её стоимость (Giddings et al., 2000). В заключение хотелось бы отметить, что несмотря на значительные достижения в области продукции реком-бинантных белков медицинского назначения в растениях, это направление находится лишь на начальном этапе своего развития. Учёные-биотехно-логи уверены, что в будущем рекомбинантные препараты, получаемые из генетически ...
0 комментариев