Навигация
1.3 СПЕЦИФИЧНОСТЬ АМИЛАЗЫ
Ферменты обладают очень высокой специфичностью. Эта специфичность обусловлена особой формой молекулы фермента, точно соответствующей форме молекулы субстрата. Эту гипотезу называют гипотезой “ключа и замка”: субстрат сравнивается в ней с ключом, который точно подходит к замку, т.е. к ферменту. Далее на основании этой гипотезы уже в 1959 году новую интерпретацию гипотезы “ключа и замка” предложил Кошланд. Он делает вывод о гибкости активных центров ферментов. Согласно этому предположению, субстрат соединяясь с ферментом, вызывает изменения в структуре последнего. Подходящей аналогией в этом случае может служить перчатка, которая при надевании на руку соответствующим образом изменяет свою форму.
Амилазы различного происхождения имеют много общих свойств: хорошо растворяются в воде или в сильно разбавленных растворах солей. Более концентрированные растворы солей (например 20-30% - ные растворы сульфата аммония) вызывают осаждение этих ферментов. Амилаза легко растворяются в разбавленных растворах этилового спирта, но осаждаются при его концентрации в среде свыше 60%. Белок амилаз обладает слабокислыми свойствами; изоэлектрическая точка ферментов колеблется в пределах рН 4,2 - 5,7. Молекулярная масса солодовой амилазы 60000, амилаз микроскопических грибов - 45000-50000. Многие из амилазы получены либо в высокоочищенном, либо в кристаллическом виде. Ионы кальция оказывают стабилизирующее действие на амилазу. Это впервые было обнаружено Воллерштейном, затем подтверждено Накамурой. В настоящее время это явление отмечено почти для всех амилаз. Однако теоретически этот вопрос применительно к промышленному гидролизу крахмала до сих пор не разработан.
Гидролиз крахмала амилазами: Амилаза действует на -1,4 -гликозидные связи, расщепляют амилозу внутри её цепи, в результате чего с большой скоростью образуются низкомолекулярные продукты гидролиза - нормальные декстрины. Их дальнейший гидролиз даёт мальтозу, мальтотриозу и глюкозу. Было найдено, что расщепление -1,4-глюкозидных связей в амилозе носит случайный характер иподчиняется закону статистического распределения продуктов реакции. Расщепление более мелких фракций на последнем этапе амилоза носит уже не случайный характер - действие фермента направлено лишь на определённые -1,4-гликозидные связи. В конечном счёте амилаза превращают амилозу в мальтозу и глюкозу, хотя и отмечены некоторые несущественные различия в динамике гидролиза этими ферментами указанного субстрата.
Бендецкий, Яровенко по изменению вязкости и восстанавливающей способности гидролизатов крахмала оценивали действие (множественность атаки) амилазы Bac. Subtilis на растворимый крахмал. Авторы наблюдали существенное различие вязкость восстанавливающая способность при кислотном и ферментативном гидролизе крахмала. Это дало основание сделать заключение, что при кислотном гидролизе деградация крахмала
Амилазы специфичны у разных видов организмов. Физиологическая роль их состоит в мобилизации запасов полисахаридов в клетках (например, при прорастании семян). Велико значение их в процессе пищеварения: Амилазы содержатся в слюне и соке поджелудочной железы человека и животных. Так, амилаза расщепляет гликоген и крахмал до глюкозы и играет важную роль во внутриклеточном обмене гликогена. Микроорганизмы потребляют крахмал, выделяя в среду амилазу.
Определение активности амилазы в сыворотке крови и моче используется в диагностике заболеваний поджелудочной железы, печени, почек, слюнных желез, а также в диагностике гликогенозов.
1.4 ВЛИЯНИЕ ИНГИБИТОРОВ И АКТИВАТОРОВ НА АКТИВНОСТЬ АМИЛАЗЫ
Действие большинства ферментов зависит от наличия ряда веществ, приводящих к инактивации ферментов. Эти вещества получили название – ингибиторов.
В основе жизненного процесса лежит обмен веществ, составными звеньями которого являются многочисленные биохимические реакции, катализируемые ферментами. Отравление (ингибирование) какого-либо одного фермента может оказать глубокое, а иногда и фатальное влияние на весь организм. Учение об ингибиторах ферментов имеет большое фундаментальное значение для фармакологии и токсикологии. Оно находит и непосредственное практическое применение, в том числе и в военном деле. Так, нервно-паралитические газы представляют собой специфические ингибиторы ферментов. В качестве инсектицидов все шире используются соединения, родственные ингибиторам ферментов. В практической медицине ингибиторы амилаз успешно применяются в качестве препаратов для лечения заболеваний, связанных с повышенной активностью этих ферментов - диабета, кариеса, ожирения и др.
В чисто академическом плане специфические ингибиторы используются как эффективные инструменты для изучения механизма действия ферментов, строения их активного центра.
Природные ингибиторы отличаются некоторыми особенностями: они действуют в чрезвычайно низких концентрациях (от 10-6 до 10-12 М), обычно являются необратимыми и т.д. Эти ингибиторы часто в природе выполняют роль регуляторов межвидовых отношений. Так, известно, что ингибиторы амилаз из проросших зерен злаковых токсичны для насекомых и млекопитающих. Недавно ингибиторы амилаз обнаружены среди так называемых "патоген - обусловленных" белков (PR-белков), участвующих в механизме защиты растений от патогенов.
Ингибиторы подразделяются на обратимые и необратимые. Необратимые ингибиторы связывают или разрушают функциональные группы молекулы фермента, необходимые для проявления его каталитической активности, при этом активность фермента не восстанавливается даже в том случае, если ингибитор удаляется каким либо способом. Необратимыми ингибиторами являются цианиды, действующие на ряд окислительно-восстановительных ферментов, содержащих в качестве кофакторов металлы. Ингибиторы типа цианидов способны образовывать устойчивые комплексы с металлами и поэтому инактивируют ферменты.
Чаще, однако, имеет место обратимое ингибирование, поддающееся количественному изучению на основе уравнения Михаэлиса-Ментен. Обратимое ингибирование в свою очередь разделяют на конкурентное и неконкурентное в зависимости от того, удается или не удается преодолеть торможение ферментативной реакции путем увеличения концентрации субстрата.
Конкурентное ингибирование может быть вызвано веществами, имеющими структуру, похожую на структуру субстрата, но несколько отличающуюся от структуры истинного субстрата. Такое ингибирование основано на связывании ингибитора с субстратсвязывающим (активным) центром. Классическим примером подобного типа ингибирования является торможение сукцинатдегидрогеназы (СДГ) малоновой кислотой. Этот фермент катализирует окисление путем дегидрирования янтарной кислоты (сукцината) в фумаровую:
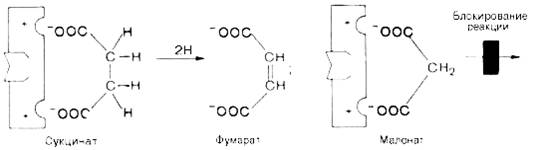
Если в среду добавить малонат (ингибитор), то в результате структурного сходства его с истинным субстратом сукцинатом (наличие двух таких же ионизированных карбоксильных групп) он будет взаимодействовать с активным центром с образованием фермент-ингибиторного комплекса, однако при этом полностью исключается перенос атома водорода от малоната. Структуры субстрата (сукцинат) и ингибитора (малонат) все же несколько различаются. Поэтому они конкурируют за связывание с активным центром, и степень торможения будет определяться соотношением концентраций малоната и сукцината, а не абсолютной концентрацией ингибитора. Таким образом, ингибитор может обратимо связываться с ферментом, образуя фермент-ингибиторный комплекс. Этот тип ингибирования иногда называют ингибированием по типу метаболического антагонизма.
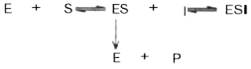
В общей форме реакция взаимодействия ингибитора с ферментом может быть представлена следующим уравнением:
![]()
Образовавшийся комплекс, называемый фермент-ингибиторным комплексом ЕI, в отличие от фермент-субстратного комплекса ES не распадается с образованием продуктов реакции. Константу диссоциации комплекса EI, или ингибиторную константу Кi, можно, следуя теории Михаэлиса–Ментен, определить как отношение констант обратной и прямой реакций:
![]()
т.е. ингибиторная константа прямо пропорциональна произведению концентрации фермента и ингибитора и обратно пропорциональна концентрации комплекса EI.
Метод конкурентного торможения нашел широкое применение в медицинской практике. Известно, например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяют сульфаниламидные препараты. Оказалось, что эти препараты имеют структурное сходство с парааминобензойной кислотой, которую бактериальная клетка использует для синтеза фолиевой кислоты, являющейся составной частью ферментов бактерий.
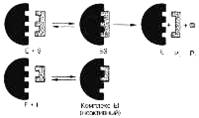
Рис. 1 Действие конкурентного ингибитора (схема по В.Л. Кретовичу). Е - фермент; S - субстрат; Р1 и Р2 - продукты реакции; I - ингибитор.
Благодаря этому структурному сходству сульфаниламид блокирует действие фермента путем вытеснения парааминобензойной кислоты из комплекса с ферментом, синтезирующим фолиевую кислоту, что ведет к торможению роста бактерий.
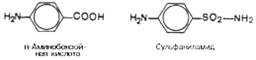
Некоторые аналоги витамина В6 и фолиевой кислоты, в частности дезоксипиридоксин и аминоптерин, действуют как конкурентные, так называемые коферментные, ингибиторы (или антивитамины), тормозящие многие интенсивно протекающие при патологии биологические процессы в организме. Применение подобных аналогов в медицинской практике (в частности, в дерматологии и онкологии) основано на конкурентном вытеснении коферментов из субстрат связывающих центров ключевых ферментов обмена.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами и часто связывающимися не с активным центром, а в другом месте молекулы фермента. Степень торможения во многих случаях определяется продолжительностью действия ингибитора на фермент. При данном типе ингибирования благодаря образованию стабильной ковалентной связи фермент часто подвергается полной инактивации, и тогда торможение становится необратимым. Примером необратимого ингибирования является действие йодацетата, ДФФ, а также диэтил-n-нитрофенилфосфата и солей синильной кислоты. Это действие заключается в связывании и выключении функциональных групп или ионов металлов и молекуле фермента.
Следует указать, что неконкурентное ингибирование также может быть обратимым и необратимым, поскольку отсутствует конкуренция между субстратом и ингибитором за активный центр. Примеры необратимого ингибирования приведены ранее. При обратимом неконкурентном ингибировании субстрат S и ингибитор I связываются с разными центрами, поэтому появляется возможность образования как комплекса EI, так и тройного комплекса EIS; последний может распадаться с освобождением продукта, но с меньшей скоростью, чем комплекс ES.
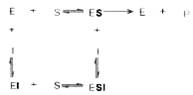
Этот тип неконкурентного ингибирования чаще всего наблюдается у ферментов, катализирующих превращения более одного субстрата, когда связывание ингибитора не блокирует связывание субстрата с активным центром. Ингибитор при этом соединяется как со свободным ферментом, так и с ES-комплексом.
Известно, кроме того, так называемое бесконкурентное ингибирование, когда ингибитор связывается с ферментом также в некаталитическом центре, однако не со свободным ферментом, а только с ES-комплексом в виде тройного комплекса.
Для выяснения вопроса о типе ингибирования пользуются уравнениями Михаэлиса-Ментен, Лайнуивера-Бэрка или другими, например уравнением Эди-Хофсти:
ν = -Km(y/[S]) + Vmax
и соответствующими графиками в прямолинейных координатах.
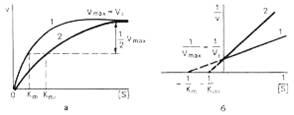
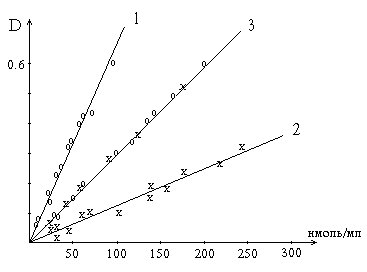
Рис. 2. Графики зависимости скорости ферментативной реакции от концентрации субстрата в присутствии конкурентного ингибитора.
а - в координатах v от [ S ] ; б - в координатах 1/v от 1 / [ S ] ; Vmaxи Vi - максимальные скорости реакции; Кm и Kmi - константа Михаэлиса соответственно в отсутствие (1) и в присутствии (2) ингибитора.
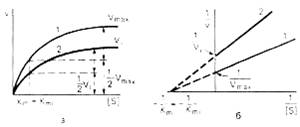
Рис. 3. Графики зависимости скорости ферментативной реакции от концентрации субстрата в присутствии неконкурентного ингибитора.
При конкурентном типе ингибирования ингибитор увеличивает значение Кm, не оказывая влияния на максимальную скорость Vmax. Это означает, что при достаточно высокой концентрации субстрата [ S ] ингибитор вытесняется молекулами субстрата из комплекса EI. При неконкурентном ингибировании (рис. 4.22) ингибитор снижает величину максимальной скорости. Если при этом величина Кm не уменьшается, то говорят о полностью неконкурентном ингибировании. Подобный тип ингибирования имеет место при образовании неактивных, труднодиссоциирующих комплексов EI и (или) EIS. Часто, однако, наблюдается смешанный тип ингибирования, иногда называемый частично неконкурентным, или обратимым неконкурентным ингибированием (см. ранее), при котором снижение Vmax сочетается с одновременным увеличением значений Кm. Это означает, что комплекс EI сохраняет частичную активность, т.е. способность к образованию промежуточного тройного комплекса EIS, в котором субстрат подвергается замедленному каталитическому превращению. В редких случаях степень торможения активности фермента может увеличиваться с повышением концентрации субстрата. Для этого типа торможения был предложен, как отмечено ранее, довольно неточный термин «бесконкурентное ингибирование». Один из механизмов такого торможения обусловлен возможностью соединения ингибитора с комплексом ES с образованием неактивного или медленно реагирующего тройного комплекса EIS.
Похожие работы
... реакция подчинена закону действующих масс и рассматривается в свете теории Михаэлиса – Ментона, то есть V=K(F) V- скорость реакции K- константа скорости F- концентрация фермента (Смотри приложение 4). На графике показано соотношение скорости реакции и концентрации субстрата. В восходящей части гиперболы при низких концентрациях ...
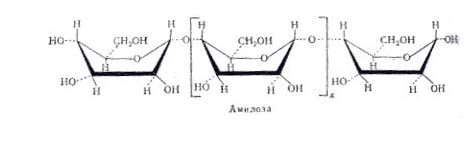
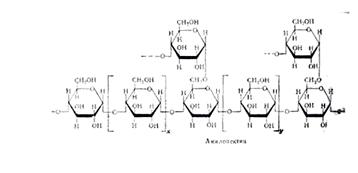
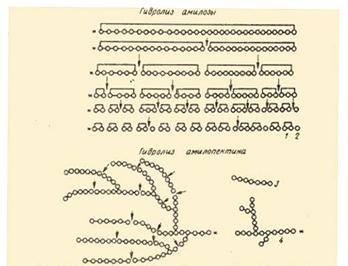
... предельным декстрином в конце гидролиза амилопектина амилазой слюны был найден тетрасахарид, солодовой амилазы – паноза, т. е устойчивым к расщеплению в первом случае были две a-1,4-глюкозидные связи, во втором – лишь одна. На рис. Представлена схема действия a-амилаз на амилозу и амилопектин по Бернфельду. Рис.2 Действие a-амилазы на амилозу и амилопектин по Бернфельду.1 – мальтоза; 2 – ...
... межпредметных связей между биологией и химией.Следует использовать знания,полученные учениками при изучении неорганической химиии в курсе 8-9 классов (темы:”Кислород,оксиды,горение”, ”Водород”, ”Кислоты,соли,основания”, “Строение вещества”). Формирование понятие “фермент” в курсе “Общая биология” Дальнейшее знакомство с ферментами ученики продолжают в курсе “Общая биология”. Здесь происходит ...
... (белка). Для целей определения ферментов могут быть использованы не только измерение поглощения света, но также измерения флюоресценции - спектрофлюорометрические методы. Такое определение активности фермента в ряде случаев по чувствительности превосходит спектрофотометрические методы на целый порядок величины. Некоторые коферменты и субстраты обладают сильной флюоресценцией. НАД и НАДФ в ...
0 комментариев