Навигация
Содержание
Введение
1.Морфоанотомические основы поглощения и движения воды
1.1 Корневая система как орган поглощения воды
1.2 Основные двигатели водного тока
1.3 Передвижение воды по растению
2.Транспирация: физиологические механизмы
2.1 Назначение транспирации
2.2 Лист как орган транспирации
3.Адаптация к дефициту воды
Заключение
Литература
ВВЕДЕНИЕ
Вода является основной составной частью растительных организмов. Её содержание доходит до 90 % от массы организмов, и она участвует прямо или косвенно во всех жизненных проявлениях. Вода- это та среда, в которой протекает все процессы обмена веществ. Она составляет основную часть цитоплазмы, поддерживает ее структуру, устойчивость входящих в состав цитоплазмы коллоидов, обеспечивает определенную конфирмацию молекул белка. Высокое содержание воды придает содержимому клетки (цитоплазме) подвижный характер. Вода – непосредственный участник многих химических реакций. Все реакции гидролиза, многочисленные окислительно-восстановительные реакции идут с участием воды.[1]
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой (тургор) обеспечивает прочность тканей, сохранение структуры травянистых растений, определенную ориентировку организмов растений в пространстве. Рост клеток в фазе растяжения идет главным образом за счет накопления воды в вакуоли.
Таким образом, вода обеспечивает протекание процессов обмена, коррелятивные взаимодействия, связь организма со средой. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органном поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой.[1]
1.Морфоанотомические основы поглощения и движения воды
1.1 Корневая система как орган поглощения воды
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Меристемы- образовательные ткани- расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля).
Определение размеров корневых систем требует специальных методов. Очень много в этом отношении достигнуто благодаря работам русских физиологов В.Г. Ротмистрова, А.П. Модестова, И.В. Красовской. Оказалось, что общая поверхность корней обычно превышает поверхность надземных органов в 104-150 раз. При выращивании одиночного растения ржи было устоновленно , что общая длинна его корней достигает 600 км., при этом на них образуется 15 млрд. корневых волосков. Эти данные говорят об огромной потенциальной способности к росту корневых систем. Однако эта способность не всегда проявляется. При росте растений в фитоценозах, с достаточно большой густотой их строение, размеры корневых систем заметно уменьшаются.[2]
С физиологической точки зрения корневая система не однородна. Довольно не вся поверхность корня участвует в поглощении волы. В каждом корне различают несколько зон(рис.1). Правда, не всегда все зоны выражены одинаково четко.
Окончание корня с наружи защищено корневым чехликом, напоминающим округлый колпачок, таящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика слущиваются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усилено делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона-зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки, затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а так же расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.[1]
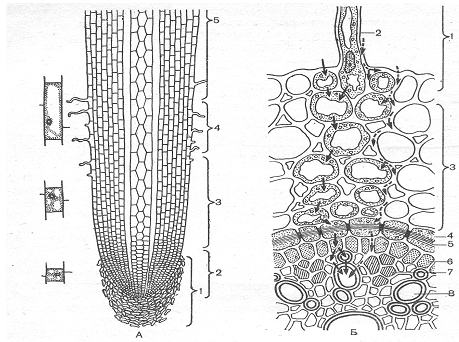
Рис.1.Схема строения корня:
А - продольный разрез: 1-корневой чехлик; 2- меристема; 3-зона растяжения; 4- зона корневых волосков; 5- зона ветвления;
Б-поперечный разрез (по М.Ф. Даниловой): 1-ризодерма; 2- корневой волосок;3-паренхима; 4- эндодерма; 5- пояски Каспари; 6- перицикл; 7- флоэма; 8-ксилема. Пунктирные стрелки- пути передвижения веществ, поглощаемых из наружного раствора. Сплошные стрелки путь растворов по симпласту ; прерывистые- путь по апопласту.
Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системных крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток- эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином. При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения, как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня. . При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление волосков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является основной зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет 5-10 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче.[1]
Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными организмами, в связи с тем что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем.
Большое значение для формирования корневых систем играет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватываются атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков.[3]
Важное значение для развития корневых систем имеет аэрация. Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания. Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений – их усиленному ветвлению.
Похожие работы
... (II) с выхлопными газами автомобилей, присутствует в выбросах свинцовых заводов и др. Ртуть – обладает повышенной возможностью распределения и биопереноса в окружающей среде. 1.3 Влияние загрязняющих веществ на морфофизиологические показатели растений Хотя сохранение растительного покрова Земли без серьезных нарушений – безусловная необходимость, его состояние в настоящее время ухудшается ( ...

... разница по отношению к контролю (Соузга, НСР05 = 0,010) несущественна для исследуемых нами сортов и форм. Некоторую разницу в общем содержании воды в листьях калины исследуемых сортов и форм можно объяснить разными погодными условиями за период вегетации (графики 3.1, 3.2): начало вегетации (конец апреля – май) характеризовался сравнительно высокими для данного периода температурами и небольшим ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев