Навигация
Основные двигатели водного тока
37099
знаков
1
таблица
3
изображения
1.2 Основные двигатели водного тока
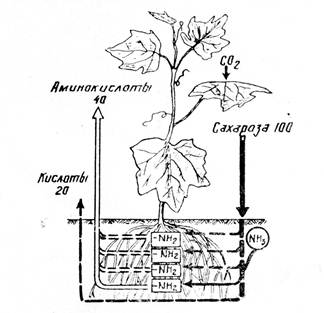
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации) , и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает) . поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.[1]
Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.
К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – эт6о не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.[1]
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
Похожие работы
... (II) с выхлопными газами автомобилей, присутствует в выбросах свинцовых заводов и др. Ртуть – обладает повышенной возможностью распределения и биопереноса в окружающей среде. 1.3 Влияние загрязняющих веществ на морфофизиологические показатели растений Хотя сохранение растительного покрова Земли без серьезных нарушений – безусловная необходимость, его состояние в настоящее время ухудшается ( ...

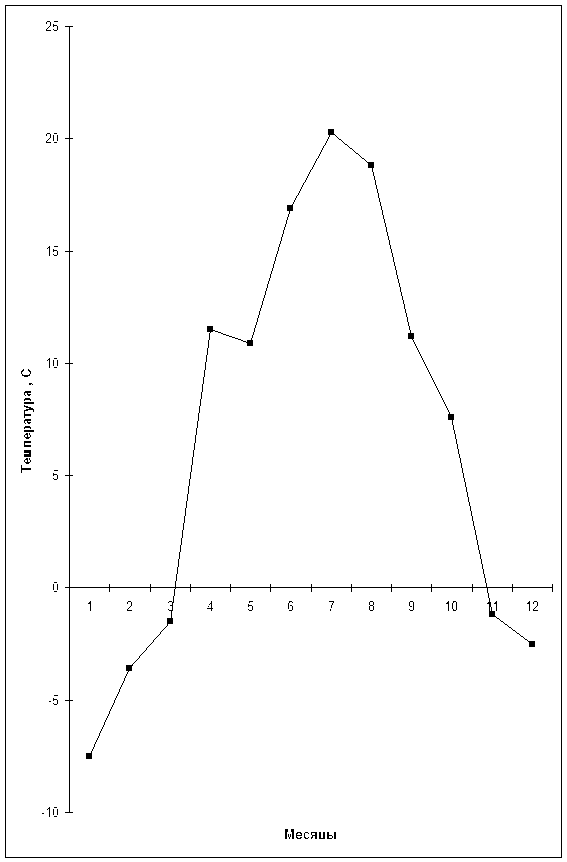
... разница по отношению к контролю (Соузга, НСР05 = 0,010) несущественна для исследуемых нами сортов и форм. Некоторую разницу в общем содержании воды в листьях калины исследуемых сортов и форм можно объяснить разными погодными условиями за период вегетации (графики 3.1, 3.2): начало вегетации (конец апреля – май) характеризовался сравнительно высокими для данного периода температурами и небольшим ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев