Навигация
Передвижение воды по растению
37099
знаков
1
таблица
3
изображения
1.3 Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.[4]
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.[5]
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 2).
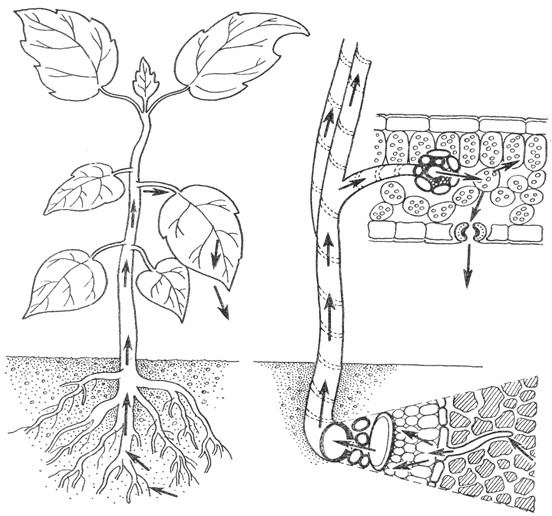
Рис.2. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.[1]
2.Транспирация: физиологические механизмы
В основе расходования воды растительным организмом лежит физиологический процесс испарения – переход воды из жидкого в парообразное состояние, происходящей при соприкосновении органов растения с не насыщенной водой атмосферой. Однако этот процесс осложнен физиологическими и анатомическими особенностями растения, и его называют транспирацией
2.1 Назначение транспирации
В обычно протекающих процессах транспирация не является необходимой. Так если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти сознательно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Известно, что большая часть всей поглощенной энергии тратится на транспирацию, которая в определенном объеме полезна растительному организму.[6]
1.Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может быть примерно 7`C ниже температуры листа завязающего, не транспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза около 30-33`C). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2.Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
3. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет процесс передвижения.[6]
2.2 Лист как орган транспирации
Основным транспортирующим органом является лист. Средняя толщина листа составляет 100-200 мкм. Паренхимных клетки листа расположены рыхло, между ними имеется система межклетников, составляющая в общей сложности от 15 до 25% объема листа. Лист окружен покровной тканью- эпидермисом, состоящим из компактно расположенных клеток, наружные стенки которых утолщены. Листья большинства растений покрыта кутикулой, в состав которой входит оксимонокарбоновые кислоты, содержащие по 16-18 атомов углерода и по 2-3 гидроксильных группы. Эти кислоты соединены друг с другом в цепочки с помощью эфирных связей. Кутикула варьирует как по составу, так и по толщине. Более развитой кутикулой характеризуются листья светолюбивых растений по сравнению с влаголюбивыми. Кутикула вместе с клетками эпидермиса образует как бы барьер на пути испарения паров воды. При этом особенно значительную преграду составляет кутикула. Удаление кутикулы во много раз повышает интенсивность испарения. Сопротивление выходу паров оказывают в определенной мере и утолщенные стенки клетки эпидермиса. Все эти особенности выработались в процессе эволюции как приспособление к сокращению испарения. Для соприкосновения листа с атмосферой имеются устьица. Устьица – одно из оригинальных приспособлений листа, обладающее способностью открываться и закрываться. Обычно устьичные отверстия ограничены двумя замыкающими клетками, стенки которых не равномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу стенки долее толстые, а внешние – более тонкие. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель. У однодольных растений строение замыкающих клеток несколько иное. Они представлены двумя удлинёнными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель. (рис.3)
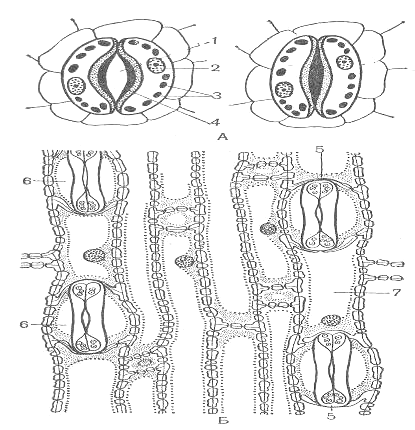
Рис.3 Структура устьиц у двудольных (А) и однодольных (Б) растений:
1-устьичная щель; 2-ядро; 3-хлоропласты; 4-толстая клеточная стенка; 5-замыкающие клетки устьиц; 6-побочные клетки; 7-клетки эпидермиса с многочисленными порами.
Число устьичных отверстий колеблется в зависимости от вида растения от 1 до 60 тыс. на 1 кв.см. листа. Большая часть устьиц расположена на нижней стороне листа. Диаметр устьичных щелей составляет всего 3-12 мкм.[1]
Устьица соединяют внутренние пространство листа с внешней средой. Вода проступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны два пути испарения: 1) через наружные стенки клеток эпидермиса в атмосферу; 2) через стенки клеток мезофилла в межклеточное пространство листа и далее в парообразном состоянии через устьица. В связи с этим различают устьичную и кутикулярную транспирацию. В том, что действительно испарение идет не только через устьица, но и через кутикулу, легко убедиться. Так, если взять листья, у которых устьица расположены только с нижней стороны ( например, листья яблони), и замазать эту сторону вазелином, то испарение воды будет продолжаться, хотя и в значительно уменьшенном размере. Следовательно, определенное количество воды испаряется через кутикулу.[7]
Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом. Однако в некоторых случаях у растений, листья которых характеризуются слабым развитием кутикулы, доля этого вида транспирации может повышаться до 30%. Имеет значение также возраст листа. Молодые листья, как правило, имеют слабо развитую кутикулу и, следовательно, более интенсивную кутикулярную транспирацию. Наименьшая Кутикулярная транспирация наблюдается у листьев, Закончивших свой рост. У старых листьев доля кутикулярной транспирации снова возрастает, так как, хотя кутикула и сохраняет достаточную толщину, в ней появляются трещины, через которые легко проходят пары воды.
Все же основная часть воды испаряется через устьица. Процесс устьичной транспирации можно подразделить на ряд этапов.
Первый этап – это переход воды из клеточных оболочек, где она находится в капельно-жидком состоянии, в межклетники (парообразное состояние). Это собственно процесс испарения. Важно подчеркнуть, что уже на этом этапе растение обладает способностью регулировать процесс транспирации (внеустьичная регулировка). Это связано с несколькими причинами: 1. Между всеми частями клетки существует водное равновесие. Чем меньше воды в клетке, тем выше становится концентрация клеточного сока. А это, в свою очередь, будет уменьшать интенсивность испарения. 2. Между микро- и макро- фибриллами целлюлозы, составляющими клеточные оболочки, имеются капиллярные промежутки. Вода испаряется именно из капилляров. Когда воды в клетках достаточно, клеточные оболочки насыщенны водой, мениски в капиллярах имеют выпуклую форму, силы поверхностного натяжения ослаблены. В этом случае молекулы воды легко отрываются и переходят в парообразное состояние, заполняя межклетники. При уменьшении содержания воды мениски в капиллярах становятся более вогнутыми, это увеличивает силы поверхностного натяжения, и вода с большей силой удерживается в клеточных оболочках. Чем более вогнут мениск, тем путь молекул воды до межклеточных пространств более длинен и извилист. В результате интенсивность испарения сокращается. Таким образом, уже на этом первом этапе растение испаряет тем меньше воды, чем меньше её содержит.[3]
Второй этап – это выход паров воды из межклетников через устьичные щели. Поверхность всех клеточных стенок, соприкасающихся с межклетными пространствами, повышает поверхность листа примерно в 10-30 раз. Все же если устьица закрыты, то все это пространство быстро насыщается парами воды и переход воды из жидкого в парообразное состояние прекращается. Иная картина наблюдается при открытых устьицах. Как только часть паров воды выедет из межклетников через устьичные щели, так сейчас же этот недостаток восполняется за счет испарения воды с поверхности клеток. Поэтому степень открытости устьиц является основным механизмом, регулирующим интенсивность транспирации. При открытых устьицах общая поверхность устьичных щелей составляет всего 1-2% от площади листа. Казалось бы, это должно очень сильно уменьшать испарение по сравнению с испарением свободной водной поверхности той же площади, что и лист. Однако это не так. Сравнение испарения листа с испарением со свободной водной поверхности той же площади показало, что оно идет не в 100 раз, как это следовало бы, исходя из размеров открытой площади (1%), а всего в два раза медленнее. Объяснение этому явлению было дано в исследованиях английских физиологов Брауна и Эскомба, которые установили, что испарение из ряда мелких отверстий идет быстрее, чем из одного крупного той же площади. Это связано с так называемым явлением краевой диффузии. При диффузии из отверстий, отстоящих друг от друга на некотором расстоянии, молекулы воды, расположенные по краям, рассеиваются быстрее. Естественно, что таких краевых молекул значительно больше в ряде мелких отверстий по сравнению с одним крупным. В связи с этим для малых отверстий интенсивность испарения пропорциональна их диаметру, а не площади. Это видно из данных таблицы.
| Диаметр пор, В мм (d) | Площадь пор, В отн.ед. (пd2) | Периметр, В отн.ед. | Испарение воды, В отн.ед. |
| 2,64 0,35 | 100 1 | 100 13 | 100 14 |
Указанная закономерность проявляется в том случае, если мелкие поры расположены достаточно далеко друг от друга. Структура листа удовлетворяет указанным требованиям. Поры (устьица) имеют малый диаметр и достаточно удалены друг от друга. При открытых устьицах выход паров воды идет достаточно интенсивно, закрытие устьиц резко тормозит испарение. Именно на этом этапе вступает в действие устьичная регулировка транспирации. При недостатке воды в листе устьица автоматически закрываются.[3]
Полное закрытие устьиц сокращает транспирацию примерно на 90%. Вместе с тем уменьшение диаметра устьичных щелей не всегда приводит к соответственному сокращению транспирационного процесса. Определения показали, что устьица должны закрываться больше чем на Ѕ, для того чтобы это сказалось на уменьшении интенсивности транспирации.
Третий этап транспирации – это диффузия паров воды от поверхности листа в более далекие слои атмосферы. Этот этап регулируется лишь условиями внешней среды.[1]
3.Адаптация к дефициту воды
Вода является необходимым условием существования всех живых организмов на Земле. Значение воды в процессах жизнедеятельности определяется тем, что она является основной средой в клетке, где осуществляются процессы метаболизма, служит важнейшим исходным, промежуточным или конечным продуктом биохимических реакций. Особая роль воды для наземных организмов (особенно растений) заключается в необходимости постоянного пополнения ее, из-за потерь при испарении. Поэтому вся эволюция наземных организмов шла в направлении приспособления к активному добыванию и экономному использованию влаги. Наконец, для многих видов растений, животных, грибов и микроорганизмов вода является непосредственной средой их обитания.[5]
Увлажненность местообитания и, как следствие, водообеснечение наземных организмов зависят, прежде всего, от количества атмосферных осадков, их распределения по временам года, наличия водоемов, уровня грунтовых вод, запасов почвенной влаги и тд. Влажность оказывает влияние на распространение растений и животных, как в пределах ограниченной территории, так и в широком географическом масштабе, определяя их зональность (смена лесов степями, степей — полупустынями и пустынями).
При изучении экологической роли воды учитывается не только количество выпадающих осадков, но и соотношение их величины и испаряемости. Области, в которых испарение превышает годовую величину суммы осадков, называются аридными (сухими, засушливыми). В аридных областях растения испытывают недостаток влаги в течение большей части вегетационного периода. В гумидпых (влажных) областях растения обеспечены водой в достаточной мере.[3]
Экологические группы растений по отношению к влаге и их адаптации к водному режиму. Высшие наземные растения, ведущие прикрепленный образ жизни, в большей степени, чем животные, зависят от обеспеченности субстрата и воздуха влагой. По приуроченности к местообитаниям с разными условиями увлажнения и по выработке соответствующих приспособлений среди наземных растений различают три основные экологические группы: гигрофиты, мезофиты и ксерофиты. Условия водоснабжения существенно влияют на их внешний облик и внутреннюю структуру.[3]
Гигрофиты — растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. Для них характерно отсутствие приспособлений, ограничивающих расход воды, и неспособность переносить даже незначительную ее потерю. Наиболее типичные гигрофиты — травянистые растения и эпифиты влажных тропических лесов и нижних ярусов сырых лесов в разных климатических зонах (чистотел большой, недотрога обыкновенная, кислица обыкновенная и др.), прибрежные виды (калужница болотная, плакун-трава, рогоз, камыш, тростник), растения сырых и влажных лугов, болот (белокрыльник болотный, сабельник болотный, вахта трехлистная, осоки), некоторые культурные растения.[3]
Характерные структурные черты гигрофитов — тонкие листовые пластинки с небольшим числом широко открытых устьиц, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие водопроводящей системы (ксилемы), тонкие слаборазветвленные корни, часто без корневых волосков. К физиологическим адаптациям гигрофитов следует отнести низкое осмотическое давление клеточного сока, незначительную водоудерживающую способность и, как следствие, высокую интенсивность транспирации, которая мало отличается от физического испарения. Избыточная влага удаляется также путем гуттации — выделения воды через специальные выделительные клетки, расположенные по краю листа. Избыточная влага затрудняет аэрацию, а следовательно, дыхание и всасывающую деятельность корней, поэтому удаление излишков влаги представляет собой борьбу растений за доступ воздуха.[5]
Ксерофиты - растения сухих местообитаний, способные переносить продолжительную засуху, оставаясь физиологически активными. Это растения пустынь, сухих степей, саванн, сухих субтропиков, песчаных дюн и сухих, сильно нагреваемых склонов. Структурные и физиологические особенности ксерофитов нацелены на преодоление постоянного или временного недостатка влаги в почве или воздухе. Решение данной проблемы осуществляется тремя способами:
1) эффективным добыванием (всасыванием) воды
2) экономным ее расходованием
3) способностью переносить большие потери воды
Интенсивное добывание воды из почвы достигается ксерофитами благодаря хорошо развитой корневой системе. По общей массе корневые системы ксерофитов примерно в 10 раз, а иногда и в 300—400 раз превышают надземные части. Длина корней может достигать 10—15 м, а у саксаула черного — 30—40 м, что позволяет растениям использовать влагу глубоких почвенных горизонтов, а в отдельных случаях и грунтовых вод. Встречаются и поверхностные, хорошо развитые корневые системы, приспособленные к поглощению скудных атмосферных осадков, орошающих лишь верхние горизонты почвы.[5]
Экономное расходование влаги ксерофитами обеспечивается тем, что листья у них мелкие, узкие, жесткие, с толстой кутикулой, с многослойным толстостенным эпидермисом, с большим количеством механических тканей, поэтому даже при большой потере воды листья не теряют упругости и тургора. Клетки листа мелкие, плотно упакованы, благодаря чему сильно сокращается внутренняя испаряющая поверхность. Кроме того, у ксерофитов повышенное осмотическое давление клеточного сока, благодаря чему они могут всасывать воду даже при больших водоотнимающих силах почвы.
К физиологическим адаптациям относится и высокая водоудерживающая способность клеток и тканей, обусловленная большой вязкостью и эластичностью цитоплазмы, значительной долей связанной воды в общем водном запасе и т. д. Это позволяет ксерофитам переносить глубокое обезвоживание тканей (до 75% всего водного запаса) без потери жизнеспособности. Кроме того, одной из биохимических основ засухоустойчивости растений является сохранение активности ферментов при глубоком обезвоживании.[5]
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев, перечисленными выше, имеют своеобразный внешний облик, за что получили название склерофиты.
К группе ксерофитов относятся и суккуленты - растения с сочными мясистыми листьями или стеблями, содержащими сильно развитую водоносную ткань. Различают листовые суккуленты (агавы, алоэ, молодило, очитки) и стеблевые, у которых листья редуцированы, а надземные части представлены мясистыми стеблями (кактусы, некоторые молочаи и др.). Фотосинтез у стеблевых суккулентов осуществляется периферическим слоем паренхимы стебля, содержащим хлорофилл. Длительные засушливые периоды преодолеваются ими путем накопления воды в водоносных тканях, связывания ее коллоидами клеток, экономного расходования, которое обеспечивается защитой эпидермиса растений восковым налетом, погруженными в ткань листа или стебля немногочисленными днем закрытыми устьицами. В результате транспирация у суккулентов чрезвычайно мала: в пустынях кактусы из рода Camegia транспирируют в сутки всего лишь I —3 мг воды на 1 г сырой массы.[3]
Корневая система поверхностная, мало развитая, рассчитана на поглощение воды из верхних слоев почвы, увлажненных редко выпадающими дождями. В засуху корни могут отмирать, но после дождей быстро (за 2—4 дня) отрастают новые. Суккуленты приурочены главным образом к засушливым зонам Центральной Америки, Южной Африки, Средиземноморья.
Мезофиты - занимают промежуточное положение между гигрофитами и ксерофитами. Они распространены в умеренно влажных зонах с умеренно теплым режимом и достаточно хорошей обеспеченностью минеральным питанием. К мезофитам относятся растения лугов, травянистого покрова лесов, лиственные деревья и кустарники из областей умеренно влажного климата, а также большинство культурных растений и сорняки. Для мезофитов характерна высокая экологическая пластичность, позволяющая им адаптироваться к меняющимся условиям внешней среды.[3]
Специфичные пути регуляции водообмена позволили растениям занять самые различные по экологическим условиям участки суши. Многообразие способов приспособления лежит, таким образом, в основе распространения растений на Земле, где дефицит влаги является одной из главных проблем экологической адаптации.[5]
Заключение
Из всего выше перечисленного можно вывести общее заключение, что при дефиците влаги растения могут адаптироваться е.т. образование корневой системы, которая достигает влажных зон почвы; ограничение расхода воды на транспирацию; запасание воды в тканях растений.
Так как вода является основной составной частью растительных организмов. Вода- это та среда, в которой протекает все процессы обмена веществ.
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой (тургор) обеспечивает прочность тканей, сохранение структуры травянистых растений, определенную ориентировку организмов растений в пространстве. Рост клеток в фазе растяжения идет главным образом за счет накопления воды в вакуоли.
Таким образом, вода обеспечивает протекание процессов обмена, коррелятивные взаимодействия, связь организма со средой. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органном поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой.
Литература
1.Якушкина Н.И. Физиология растений. М., Просвещение, 1980г.
2.Козловский Водный обмен растений. М., Колос. 1969г.
3.Сказкин Ф.Д. Критический период у растений к недостаточному водоснабжению. М., Наука. 1968г.
4.Радкевич В.А. Экология. Мин., Высшая школа. 1983г.
5.Генкель П.А. Физиология устойчивости растительных организмов. М., Изд-во МГУ. 1967г. т.3.
6. Жданов В.С. Аквариумные растения. / Под. ред. Коровина. -
М.: Лесн. пром-ть , 1981
7. Горышина Т.К. Экология растений. – М.: Высшая школа,1979
Похожие работы
... (II) с выхлопными газами автомобилей, присутствует в выбросах свинцовых заводов и др. Ртуть – обладает повышенной возможностью распределения и биопереноса в окружающей среде. 1.3 Влияние загрязняющих веществ на морфофизиологические показатели растений Хотя сохранение растительного покрова Земли без серьезных нарушений – безусловная необходимость, его состояние в настоящее время ухудшается ( ...

... разница по отношению к контролю (Соузга, НСР05 = 0,010) несущественна для исследуемых нами сортов и форм. Некоторую разницу в общем содержании воды в листьях калины исследуемых сортов и форм можно объяснить разными погодными условиями за период вегетации (графики 3.1, 3.2): начало вегетации (конец апреля – май) характеризовался сравнительно высокими для данного периода температурами и небольшим ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев