Навигация
Ембріональний розвиток ссавців
51123
знака
0
таблиц
0
изображений
2. Ембріональний розвиток ссавців
2.1 Доімплантаційний розвиток
У ссавців живлення за рахунок яйцеклітин відбувається тільки на ранніх стадіях, а далі, навіть до народження, воно здійснюється виключно за рахунок організму матері. Яйцеклітини вкриті двома оболонками – первинною і вторинною. Первинна – це плазмолемма клітини, вторинна утворюється фоллікулярними клітинами, з яких побудовані стінки фоллікула і які оточують яйцеклітину після її виходу з яєчника [2].
Дроблення у різних тварин відбувається по-різному. Процес дроблення великої рогатої худоби триває 8 днів, з яких 4 дні – в яйцепроводі і 4 дні в матці. У щурів доімплантаційний період складає 6 днів. Повне дроблення ссавців відбувається несинхронне - в результаті утворюються бластомери різної величини і зародок відповідно складається з 3,5,7 і т.д. бластомерів. Останні звичайно лежать у вигляді купки клітин (ця стадія носить назву морули). Вже у процесі дроблення відбувається відокремлення зародкових частин від незародкових – бластомери утворюються двох типів: дрібні світлі і більш грубі темні. [1]
Дрібні і світлі бластомери розташовуються ззовні і утворюють трофобласт, поступово обростаючи більш грубими темними бластомерами. Трофобласт в подальшому не приймає участі у побудові тіла зародка, але відіграє велику роль при імплантації та подальшому живленні ембріона.
Грубі і темні клітини утворюють ембріобласт, за рахунок якого формується тіло зародка, а пізніше виникають всі зовнішньозародкові органи.
Дроблячись, зародок пересувається по яйцепроводу у напрямку матки і поглинає секрет маткових залоз, який накопичується між трофо- і ембріобластом. Клітини трофобласту поглинають рідину і формують порожнину, розміри якої швидко збільшуються - утворюється трофобластичний пухирець. Він складається із зовнішнього шару клітин – трофобласту і внутрішнього скупчення клітин - ембріобласту, які, прилягаючи до трофобласту формують зародковий вузлик та порожнину бластули, яка заповнена рідиною. Ця стадія носить назву бластоцисти. [4]
У корів такий пухирець розвивається впродовж 7-11 дів і має форму кульки. На 12-ту добу діаметр пухирця у корів становить 1 мм, а на 13-ту він видовжується і становиться веретеноподібним.
Розвиток бластули відбувається під прикриттям яскравої оболонки колишньої яйцеклітини і третинними оболонками. Після вилуплення (розрив яскравої оболонки і вихід бластули) бластула імплантується в слизову оболонку матки. Клітини трофобласту синтезують специфічні ферменти, які руйнують тканину матки і зародковий пухирець, занурюючись в них, контактує з організмом матері. У корови це відбувається на 13-15-й день, у щурів на 7-й день після заліднення [17].
2.2 Загальна схема імплантації
Імплантація ембріона у порожнину матки – складний, багатоступінчастий процес, регуляція якого здійснюється за участі великої кількості гуморальних факторів і різноманітних міжмолекулярних та міжклітинних взаємодій.
Як було зазначено вище, запліднена яйцеклітина потрапляє у порожнину матки на стадії морули. Там вона розвивається у бластоцисту. Материнській організм сприйнятливий до бластоцисти, що імплантується тільки у межах строго визначеного по часу “вікна імплантації”. Поява у ендометрії трансмембранного глікопротеїну MUC-1 обмежує часові рамки “вікна імплантації” [27].
Під час фаз опозиції (протистояння ембріона і матки) та прикріплення на зовнішній мембрані бластоцисти утворюються багаточисельні мікровип’ячування – піноподи, в результаті чого вона входить у тісний контакт з матковим епітелієм, що означає перехід в стадію адгезії. Від’ємний електричний заряд на поверхні епітеліальних клітин сприяє зближенню бластоцисти з поверхнею ендометрію [25].
2.3 Роль стероїдних гормонів в імплантаційних процесах
Основними регуляторами морфологічних змін функціонального шару ендометрію протягом статевого циклу та у доімплантаційний період є стероїди, що синтезуються яєчниками. Тільки підготовлений циклічним впливом стероїдних гормонів ендометрій буде готовим до сприйняття бластоцисти та її гуморальних сигналів [13].
Встановлено, що ембріони самі можуть впливати на функції материнського організму протягом періоду імплантації. До імплантації розвиток ембріона залежить від секретів яйцепроводів і матки, від особливостей будови і функцій статевих органів самок, в яких розвивається бластоциста. Доказом цього є те, що культивування ембріонів корів до стадії бластоцисти можливо лише у поживному середовищі, максимально наближеним за хімічним складом до середовища яйцепроводів [22].
Відомо, що материнські стероїдні гормони регулюють синтез гормонів статевого тракту і таким чином можуть впливати на розвиток ембріона. Ембріональний розвиток після стадії одношарової бластоцисти відбувається нерівномірно до тих пір, поки ембріон не звільняється від зони пеллюцида – це відбувається під дією протеолітичних ферментів, які утворюються в ендометрії. Ці ферменти контролюються стероїдами яєчників, а їх активність стимулюється самою бластоцистою та починається на початкових етапах імплантації
Для всіх видів ссавців під час статевого циклу характерні дві слідуючі закономірності:
· зростаюча секреція естрогенів у фазі селекції і розвитку фолікулів, яка у хронологічному відношенні тільки неопосередкованно впливає на імплантацію;
· синтез великих кількостей прогестинів у секреторну фазу циклу, що співпадає по часу з імплантацією.
Рахується, що на відміну від прогестерону, естрогени впливають на імплантацію опосередковано. Естрадіол виступає у ролі пермісивного агента, тоді як прямий вплив характерний для локальних факторів, яких він регулює – цитокінів, молекул адгезії і факторів росту [23]. Більше того, вирішальну роль в імплантації відіграє не абсолютний вміст стероїдних гормонів, які діють на тканини-мішені органів репродуктивної системи, а морфологічна структура ендометрію, а саме ступінь рецептивності останнього, тобто кількість функціонально повноцінних рецепторів до відповідних стероїдних гормонів. [5]
Естрогени, одночасно із стимуляцією проліферації клітин маткового епітелію, активують розвиток секреторного апарату клітин ендометрію і синтез рецепторів до естрогенів та прогестерону, за допомогою яких і здійснюється різноманітний вплив гормонів на клітини. Вся родина стероїдних рецепторів являє собою клас білків, що функціонують на ядерному рівні і по суті, регулюють транскрипційні процеси. У відсутність лігандів стероїдні рецептори знаходяться у комплексі з білками теплового шоку. При зв’язуванні з стероїдом відбувається дисоціація цього комплексу і димеризація рецептора. Димер володіє здатністю зв’язуватись з відповідними, строго специфічними для стероїдів послідовностями ядерної ДНК, що забезпечує селективність у відношенні кожного з них і впливає на транскрипцію генів-мішеней. Всі стероїдні рецептори можуть конкурувати за одні і ті ж загальні транскрипційні факторі.
Більша частина гормон-рецепторних комплексів у ядрі дисоціюється і інактивується. В цитоплазму повертаються вільний стероїд та інактивований під дією ядерних фосфатаз рецептор. Гормон ніяких змін не зазнає.
Головним фізіологічним регулятором експресії ядерних рецепторів всередині клітин-мішеней є рівень циркулюючих вільних гормонів. Естрадіол підсилює синтез власних рецепторів, рецепторів до прогестерону і рецепторів андрогенів. Прогестерон, навпаки, пригнічує синтез як власних рецепторів, так і рецепторів естрадіолу [5], [14].
.
3. Фізіологічні та молекулярні механізми імплантаційних процесів
3.1 Модель імплантації у ссавців
Багаточисельні дослідження на модельних тваринах, головним чином на мишах, виявили велику кількість молекул, які приймають участь у процесах імплантації. Генетичні дослідження за допомогою targeting-методики допомогли ідентифікувати багато факторів росту та їх рецепторів, включаючи епідермальний фактор росту (ЕФР), трансформуючі фактори росту (ТФР) та інтерлейкіни, які приймають участь у процесах імплантації ембріонів. Крім того, in vitro дослідження показали, що гепарин-зв’язаний епідермальний фактор росту стимулює адгезію бластоцистів миші, а фібронектин та його рецептори задіяні у процесах утворення відростків трофобласту при адгезії бластоцист. [9]
In vitro дослідження імплантації у людини виявили наступні стадії:
Похожие работы
... кількості мітохондрій в період оогенезу. У багатьох тварин розмноження мітохондрій відбувається в основному до початку вітелогенезу. Наприклад, у жаби в період вітелогенезу і на ранніх стадіях ембріогенезу мітохондрії практично не діляться, тобто інтенсивне розмноження іде в превітелогенних ооцитах [Айзенштадт, 1984]. Таким чином, активна реплікація мітохондріальної ДНК в превітелогенних ооцитах ...
... впливу фосфорорганічного пестициду діазинону дозоволило авторам роботи зробити висновок про значні морфологічні та фізіологічні зміни у Melanotaenia duboulayi, що виражались у гіпертрофії та гіперплазії зябер, закупорці кровоносних судин, потовщенні епітелію зябрових пелюсток, посиленні секреції слизу. При хронічному отруєнні похідними дихлорфеноксиоцтової кислоти у риб спостерігається ...
... ів України. – Київ, 2003. – С.312. (Внесок дисертанта: проведено визначення каталази в крові хворих і підготовлено матеріал до друку) АНОТАЦІЯ Поліянц І.В. Патофізіологічні механізми пневмонії на різних етапах її розвитку. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за фахом 14.03.04 – патологічна фізіологія. Одеський державний медичний університет МОЗ Укра ...
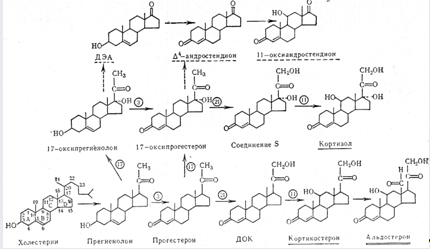


... контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою. Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо. Кіркова і ...
0 комментариев