Навигация
Людська вилуплена бластоциста прикріплюється, а потім проростає у моношар внутрішньоматкової строми;
51123
знака
0
таблиц
0
изображений
1. Людська вилуплена бластоциста прикріплюється, а потім проростає у моношар внутрішньоматкової строми;
2. Трофобласт розпластовуєтьсяя по всій поверхні моношару стромальних клітин та з двох полюсів проростає в ці клітини;
Спільне культивування бластоцист та культури стромальних клітин значно стимулює синтез бластоцистами hCG. [7]
Таким чином, імплантація у всіх ссавців може бути описана як трьохстадійний процес: спочатку бластоциста активує відповідні ферментативні перетворення в ендометрію матки, які будуть сприяти процесам імплантації – це досягається двостороннім паракринним “діалогом” між матковою оболонкою і бластоцистою. Далі трофоектодерма приєднюється до люмінального епітелію оболонки матки шляхом взаємодії сигнальних молекул та їх рецепторів, і таким чином проростає через шар епітеліоцитів до базальної мембрани. І остання, третя стадія полягає у проростанні трофоектодерми в основну строму матки. Всі епітеліальні клітини синтезують інтегрини 3, 1 і 4, лептин та глікопротеїд MUC-1 – епітеліальне похідне глікокаліксу. Вважається, що саме його нестача на поверхні місця прикріплення бластоцисти і сповільнює імплантаційні процеси [20].
3.2 Функціональні зміни у тканинах матки при імплантації
Маточний ендометрій протягом доімплантаційного періоду багаторазово змінюється. Всі його компоненти - залозистий епітелій, покривний епітелій, стромальні клітини та міжклітинний матрикс зазнають морфологічних, клітинних і молекулярних змін [20], [25].
Залозистий епітелій. В період імплантації збільшується мітотична активність залоз. Секреторна активність залоз сягає максимума. Плацентарний протеїн 14 (ПП14) - головний компонент секреторного продукту залоз ендометрію протягом другою половини лютеїнової фази циклу і ранніх стадій вагітності. Це глікопротеїн з Мм 42 кД – прогестеронзалежний ендометріальний білок, який попереджує лімфопроліферацію і пригнічує активність природних клітин-кіллерів, яких є особливо багато на ранніх стадіях вагітності. Цей білок захищає ембріон, що імплантується від імунної системи матері.
Покривний епітелій. Це тканина, яка вкриває внутрішню порожнину матки. Він першим контактує з бластоцистою і саме в ньому відбуваються основні анатомічні і молекулярні зміни, які підвищують рецепторну активність ендометрію до нідації бластоцисти. У період імплантації на поверхні покривного епітелію з’вляються піноподи, цей процес інгібуться естрогеном і активуються прогестероном, їх поява співпадає з утворенням “вікна імплантації” – точки найвищої рецептивної активності ендометрію. MUC-1 – циклозалежний глікопротеїн, який був виявлений у мишей і людини. У мишей синтез мРНК MUC-1 та її білкового продукту в покривному епітелії зменшується в період діеструсу і стає майже непомітним перед адгезією бластоцисти. У людини синтез MUC-1 в ендометрії сягає максимума саме під час імплантації, що доводить двояку його роль у різних видів ссавців.
Стромальні клітини складаються, головним чином, з 2-х різних популяцій клітин: фібробластів і лейкоцитів. У другій половині секреторної фази статевого циклу фібробласти під дією прогестерону трансформуються у псевдодецидуальні клітини. У цьому процесі приймають участь такі регуляторні молекули, як гістаміни, простагландини, лейкотрієни, плацента-стимулюючий фактор, інтерлейкін-1 і ангіотензин. Ці механізми відповідають за контроль неадекватної імуносупресії, коли організм матері починає відповідати на вростання трофобласта негативною імунною реакцією.
Зовнішньоклітинний матрикс. Більшість його компонентів виробляється стромальними фібробластоми. Протягом проліферативної фази статевого циклу фібронектин і колагени типів ІІІ, V та VI є головними інтерстеціальними компонентами. При наближенні імплантації колагенові волокна розсмоктуються і міжклітинний матрикс стає менш в’язким, знижується стромальна імунореактивність до колагенів типів ІІІ і V, а колаген типу VI майже зовсім зникає – це спричиняє набряк слизової та збільшення зовнішньоклітинного простору. На ранній стадії лютеїнової фази циклу навкого стромальних клітин з’являються ламінін та фібронектин. Колаген починає активно синтезуватись на заключній стадії циклу – на початку трансформації стромальних клітин у перидецидуальні. Рахується, що саме зовнішньоклітинний матрикс чітко визначає межи, у яких відбуваються клітинні процеси раннього ембріонального розвитку: міграція, імплантація, а потім і плацентація. [27]
Таким чином, процес імплантації контролюється складними взаємодіями великої кількості сигнальних і ефекторних сполук, які продукуються ендометрієм, імунокомпетентними клітинами матері та ембріоном. Однак власно імплантації передують процеси, які розвиваються в ендометрії в секреторній фазі статевого циклу. У відповідності з цим реакція ендометрію в ході імплантації поділяється на три фази:
Перша фаза. Знаходиться під контролем естрогенів та прогестерона і характеризується змінами у покривних та залозистих епітеліальних клітинах ендометрію, результатом чого є підготовка до опозиції та приєднання бластоцисти. Гормональні впливи на ендометрій залежать від наявності ядерних рецепторів до стероїдних гормонів. Наростання концентрації рецепторів спостерігається у напрямі від функціонального шару до базального, що корелює із встановленням максимальної рецептивності матки, необхідної для імплантації. Паралельно зі змінами в системі стероїдних рецепторів, клітини епітелію піддаються змінам у структурі цитоскелету та профілю секреції білків. Ці зміни можуть бути попереджені антагоністами прогестеронових рецепторів у лютеїновій фазі циклу.
Друга фаза. Модуляція гормональних стероїдних ефектів ембріональними факторами. Початок секреції бластоцистою хоріонічного гонадотропіну та інших білків ранньої вагітності викликає додаткові зміни у клітинах ендометрію. В клітинах покривного епітелію відбувається ендореплікація, утворюються “епітеліальні бляшки”. Залозистий епітелій відповідає на дію ембріональних регуляторів модифікацією головного секреторного продукта – глікоделіну, який дає імуннопротекторний ефект у відношенні вагітності, що починається. Стромальні фібробласти починають процес свого диференціювання, набувають децидуального фенотипу и починають продукувати актинові філаменти.
Третя фаза. Інвазія трофобласта і перебудова стромального компонента ендометрію. При цьому на покривному епітелії зникають “епітеліальні бляшки”, залозистий епітелій залишається високо секреторно активним. В цій фазі закінчується трансформація фібробластів в децидуальні клітини, які починають синтезувати і секретувати весь спектр ростових факторів, які властиві для ранніх стадій вагітності.
Похожие работы
... кількості мітохондрій в період оогенезу. У багатьох тварин розмноження мітохондрій відбувається в основному до початку вітелогенезу. Наприклад, у жаби в період вітелогенезу і на ранніх стадіях ембріогенезу мітохондрії практично не діляться, тобто інтенсивне розмноження іде в превітелогенних ооцитах [Айзенштадт, 1984]. Таким чином, активна реплікація мітохондріальної ДНК в превітелогенних ооцитах ...
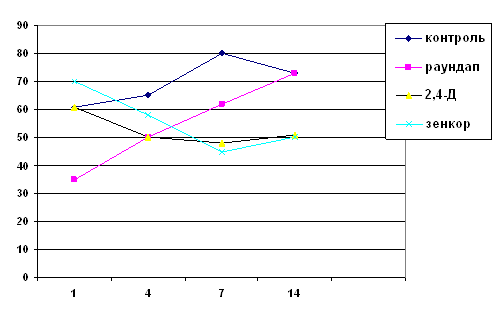
... впливу фосфорорганічного пестициду діазинону дозоволило авторам роботи зробити висновок про значні морфологічні та фізіологічні зміни у Melanotaenia duboulayi, що виражались у гіпертрофії та гіперплазії зябер, закупорці кровоносних судин, потовщенні епітелію зябрових пелюсток, посиленні секреції слизу. При хронічному отруєнні похідними дихлорфеноксиоцтової кислоти у риб спостерігається ...
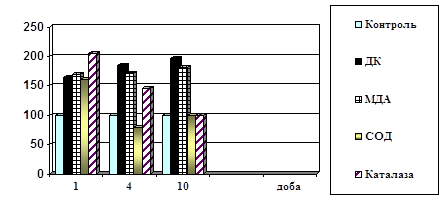
... ів України. – Київ, 2003. – С.312. (Внесок дисертанта: проведено визначення каталази в крові хворих і підготовлено матеріал до друку) АНОТАЦІЯ Поліянц І.В. Патофізіологічні механізми пневмонії на різних етапах її розвитку. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за фахом 14.03.04 – патологічна фізіологія. Одеський державний медичний університет МОЗ Укра ...
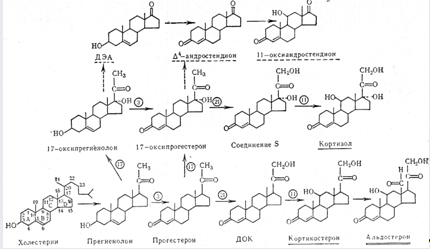

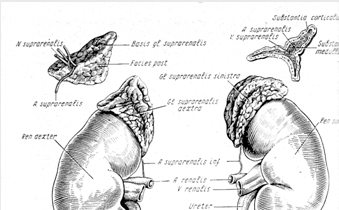

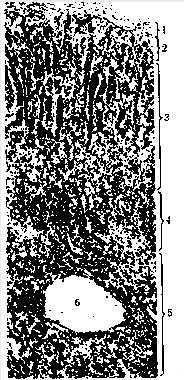
... контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою. Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо. Кіркова і ...
0 комментариев