Навигация
Механізми дії ростових факторів
51123
знака
0
таблиц
0
изображений
4.2 Механізми дії ростових факторів
На відміну від ендокринних механізмів регуляції клітинної проліферації існують паракринні механізми, які діють на тканинному рівні. При цьому ростові фактори здійснюють дію головним чином на сусідні клітини.
Аутокринний тип регуляції – це коли клітини відповідають на дію фактору, який продукують самі і мають на нього рецептори. Цей тип регуляції має велике значення в період раннього ембріонального розвитку, коли йде активне розмноження ембріональних плюрипотентних клітин.
На відміну від стероїдних гормонів, які синтезуються з одного загального попередника, кожен поліпептидний фактор росту утворюється із свого попередника в результаті його процесінгу. Є дані, що ці попередники – трансмембранні білки, а активна форма фактору росту утворюється в результаті протеолітичного розщеплення зовнішньоклітинної ділянки молекули-попередника – тобто попередник є поліфункціональним пепетидом, а сама трансмембранна форма деяких факторів є рецептором для ліганду, що входить у його склад. Таким чином, є підстава вважати, що попередники деяких ростових факторів проявляють біологічну активність, залишаючись вбудованими в плазматичну мембрану клітин – у такому випадку вони взаємодіють із специфічними рецепторами сусідніх клітин без попереднього процесінгу. Такий спосіб регуляції клітинної проліферації називається юкстакринним. Є данні, що юкстакринна регуляція відіграє роль не тільки при міжклітинній адгезії, але і у ході стимуляції проліферації клітин. Особливо важливим це є в періоди раннього ембріонального розвитку, коли ще відсутні клітини, які секретують спеціальні гормони та ростові фактори [4], [8].
У послідовності взаємодії ростового фактора та клітини-мішені можна виділити певні послідовності:
· взаємодія фактору росту із специфічним рецептором плазматичної мембрани клітини-мішені;
· утворення і активація ліганд-рецепторного комплексу;
· біологічні ефекти, які виникають під дією регуляторних сигналів, індукованих фактором росту.
4.3 Взаємодія рецепторів факторів росту з лігандами
Рецептори факторів росту розташовані на поверхні зовнішньої мембрани клітин. Вони представлені численним класом глікопротеїдів з достатньо великою молекулярною вагою. Найбільш детально вивчено будову рецепторів інсуліну та епідермального фактор росту (ЕФР). Рецептор інсуліну складається з двох α- та двох β-субодиниць, які з’єднані між собою дисульфідними містками. Молекула рецептора складається із трьох основних функціонально різних доменів: ліганд-зв’язуючого, який багатий на вуглеводневі структури, трансмембранного, який складається переважно із залишків гідрофобних амінокислот, і цитоплазматичного, якій володіє ендогенною протеінкназною активністю. Цьому домену, який каталізує фосфорилювання білкових субстратів плазматичної мембрани і цитозолю по залишках тирозину, відводиться важлива роль у механізмах передачі регуляторного сигналу всередину клітин-мішеней. [12], [3]
Зв’язування рецептора з фактором росту забезпечує димеризацію і активацію тирозинкінази. Так ініціюється каскад реакцій фосфорилювання. Субстратами фосфорилювання є різні кінази, фосфоліпази і самі рецептори факторів росту. В якості біологічних ефектів фосфорилювання відбувається зміна мітогенних характеристик тканин, і диференціювання клітин, стимуляція транспортних систем і активація метаболічних шляхів.
Після взаємодії фактора росту з рецепторами клітинної поверхні відбувається процес кластеризації ліганд-рецепторних комплексів – процес їх агрегації у визначених місцях клітинної поверхні. Це необхідно для подальшого проникнення комплексів всередину клітин-мішеней. Після взаємодії інтенсивність зв’язування лігандів на поверхні клітин значно знижується – частина ліганд-рецепторних комплексів інтерналізується (піддається розщепленню в лізосомах), а частина повертається на поверхню клітин. Також виявлено, що після впливу деяких факторів росту на клітини у останніх змінюється здатність до зв’язування інших факторів росту.
Утворення комплексів факторів росту з рецепторами призводить до значних змін останніх. На відміну від рецепторів стероїдних гормонів, рівень мРНК рецепторів факторів росту багатократно збільшується протягом декількох годин після впливу. [2], [3].
4.4 Фактори росту на різних стадіях ембріонального розвитку
Всі періоди розвитку ембріонів співпадають у часі з перебудовою їх білкового складу. Перехід з однієї стадії в іншу пов’язують з регуляторним впливом специфічних білків, які діють на клітини локально по аутокринному або паракринному механізму. Під час аутокринної регуляції клітини, які секретують ростовий фактор, є одночасно і мішенями його дії, а під час паракринної регуляції фактори росту діють на сусідні клітини - мишені. За допомогою цих механізмів поліпептидні фактори росту виконують регуляторну роль в ембріональному періоді розвитку тварин.
Паракринні механізми регуляції клітинної проліферації, що діють на тканинному рівні, є еволюційно більш давніми ніж ендокринні. За цих умов фактори росту діють на клітини зблизька. Вони секретуються у позаклітинний простір, де можуть зв’язуватися зі специфічними рецепторами, що знаходяться на інших клітинах і які синтезують цей фактор росту [3].
Довгий час лишалося невідомим чи є аутокринна регуляція проліферації клітин феноменом, що характерний тільки для клітинної трансформації, чи це є загальнобіологічний механізм, за допомогою якого регулюються процеси клітинного розмноження. Згодом дослідженнями на ембріональних клітинах ссавців було доведено, що аутокринна регуляція проліферації має важливе значення під час ранніх періодів розвитку, коли відбувається активний поділ первинних ембріональних клітин [2].
Вже на початку доімплантаційного періоду ряд факторів росту починає проявляти ембріотрофічну активність. Наявність ТФР-α і внутрішньоклітинного домену до рецептора ЕФР вже виявляється у 4-клітиного ембріона. У 8- та 14-клітинних ембріонів виявляються ТФР- α, внутрішньо- та зовнішньоклітинні домени до рецепторів ЕФР, ІФР-1 та його рецептор. ТФР-β виявляється у ембріонах вівці на 12-16-й день, тоді як в культурі ендометрію його взагалі не має. Припускають, що продукування ембріоном ТФР-β розпочинається після материнського “впізнання” вагітності і співпадає по часу з імплантацією. Доведено, що родина ТФР-β відіграє важливу роль в цих процесах, адже саме вони забезпечує рецептивність ендометрію у відношенні бластоцисти. Децидуальний ТФР-β материнського походження може пригнічувати надлишкову інвазію трофобласта, а введення антагоністів ТФР-β викликало повне інгібування імплантації бластоцисти.
В останні роки багато уваги приділяється вивченню юкстакринної регуляції через ТФР-α в клітинах людського ендометрію при імплантації. Показано, що трансмембранна форма ТФР-α епітелію матки впливала на рецептор ЕФР мишиної бластоцисти при прямому міжклітинному контакті. Індукція та пік синтезу ТФР-α у мембранах епітелію матки співпадали по часу з періодом імплантації та були пов’язані з відповідним зростанням експресії рецепторів ЕФР на бластоцисті. При зв’язуванні ТФР-α з рецептором ЕФР розвивається два види відповіді: мітогенна і активація внутрішньоклітинних сигнальних шляхів. Така двостороння дія має дуже важливе значення у репродукції – при експресії ембріоном рецепторів ЕФР, які можуть взаємодіяти з ТФР-α ендометрію материнського організму, з одного боку стимулюється ріст і диференціювання ембріона, а з іншого – активується мітогенний сигнал у самих клітинах ендометрію, де відбувається інвазія. [10], [8].
Ранні доімплантаційні ембріони мишок і корів мають рецептори різних ростових факторів серед яких трансформуючий ростовий фактор TGF-α і TGF-β та інсуліноподібний ростовий фактор IGF-І, що в більшості випадків починають проявлятися на стадії незаплідненої яйцеклітини [19]. Доведено, що епідермальний ростовий фактор (EGF) здатний стимулювати ембріональний розвиток мишачих ембріонів у культурі. Інсулін стимулював синтез білків у 8-ми клітинних ембріонах мишок, бластоцистах і експандованих бластоцистах, специфічно впливав на диференціацію, оскільки він стимулював клітинне дроблення тільки внутрішньої клітинної маси, в той час як трофоектодермальні клітини лишалися „байдужими” до цього впливу [15]. Вплив інсуліну на ембріональний розвиток був очевидним навіть до 8-бластомерної стадії, коли виявляли тільки м-РНК інсулінового рецептору і не було виявлено будь-яких білків [21].
Рецептори IGF-1 було виявлено в ранньому ембріональному розвитку і доведено, що IGF-1 активує стимуляцію ембріонального метаболізму починаючи з 2-клітинної стадії розвитку. Попередніми дослідженнями на ембріонах свиней [8] виявлено вплив інсуліну на біосинтез естрадіолу, що дуже важливо для маткових шляхів, стимулює ембріональний диск до синтезу чотирьох нових білків, які потім беруть участь у механізмі материнського розпізнавання вагітності [24]. В цьому контексті ідентифікація IGF-І та IGF-II у рідині яйцепроводів свині разом з присутністю у епітелії яйцепроводів корів IGF-ІІ транскрипцій дозволило припустити, що білки, які пов’язані з інсуліном, є медіаторами впливу яйцепроводів на ембріональний розвиток. Відмічають, що яйцепроводи овець і корів секретують білки, які посилюють дію інсуліну в стимуляції мітозів ембріонального трофобласту [24].
Паралельні дослідження довели присутність м-РНК TGF-α та TGF-β, епідермального фактора росту, фактора росту тромбоцитів PDGF та інших ростових факторів у яйцепроводах ссавців [3]. Фактори росту PDGF, активін та епідермальний ростовий фактор ідентифіковано серед білків, які секретуються і локалізуються в окремих ділянках протоків яйцепроводів [25].
Зустрічається застереження щодо дії високих концентрацій ростових факторів у середовищі, які пригнічують проліферацію клітин яйцепроводів та кумулюсу [22].
Експерименти ряду авторів з вивчення впливу факторів росту на розвиток і дроблення ранніх ембріонів тварин доводять позитивний ефект цих поліпептидів при культивуванні цих ембріонів in vitro на фідерних моношарах. Y. Tsuchiya et al. підтвердили суттєвий вплив EGF в кількості 10нг/мл, інсуліну- 20мкг/мл не тільки на розвиток in vitro бластоцист овець, але й змогли отримати з їх внутрішньої клітинної маси колонію стовбурових ембріональних клітин, які були життєздатними на протязі 8 тижнів. Цікаво, що інсулін більш сприятливо впливав на формування ЕСК-колоній. Створені колонії клітин росли добре, але після першого пасажу зменшили інтенсивність росту, зберігаючи свою морфологію. Додавання EGF у середовище також спричинило ріст деяких колоній, але колонії були маленькі і росли слабо. Після двох пасажів вони також зберігали свою морфологію.
За даними Y. Iwakura et al присутність лейкеміє інгибуючого ростового фактору (LIF) та EGF в кількості 10 нг/мл сприяло суттєвому збільшенню колонії стовбуроподібних (плюріпотентних) клітин при культивуванні їх на фідерному моношарі ембріонального фібробласта протягом 5 днів. Подібні результати були раніше отримані на вівцях із застосуванням двох типів фідерних шарів: стовбурових клітин, що сформовані плюріпотентними клітинами бластоцист мишок і ембріональним фібробластом, які одержали з плодів вівцематок 32-го дня вагітності [17].
Glister C. at el в експерименті спостерігали in vitro дозрівання і запліднення 1761 ооцитів, дійшли висновку, що додавання ростових факторів, таких як IGF-1, TGF-a, EGF та PDGF в кількості 10 нг/мл до культурального середовища, однак, сприяє швидкості дроблення ембріонів, розвитку до стадії бластоцисти і вилуплення бластоцист з прозорої оболонки в присутності сироватки. [15]
Похожие работы
... кількості мітохондрій в період оогенезу. У багатьох тварин розмноження мітохондрій відбувається в основному до початку вітелогенезу. Наприклад, у жаби в період вітелогенезу і на ранніх стадіях ембріогенезу мітохондрії практично не діляться, тобто інтенсивне розмноження іде в превітелогенних ооцитах [Айзенштадт, 1984]. Таким чином, активна реплікація мітохондріальної ДНК в превітелогенних ооцитах ...
... впливу фосфорорганічного пестициду діазинону дозоволило авторам роботи зробити висновок про значні морфологічні та фізіологічні зміни у Melanotaenia duboulayi, що виражались у гіпертрофії та гіперплазії зябер, закупорці кровоносних судин, потовщенні епітелію зябрових пелюсток, посиленні секреції слизу. При хронічному отруєнні похідними дихлорфеноксиоцтової кислоти у риб спостерігається ...
... ів України. – Київ, 2003. – С.312. (Внесок дисертанта: проведено визначення каталази в крові хворих і підготовлено матеріал до друку) АНОТАЦІЯ Поліянц І.В. Патофізіологічні механізми пневмонії на різних етапах її розвитку. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за фахом 14.03.04 – патологічна фізіологія. Одеський державний медичний університет МОЗ Укра ...
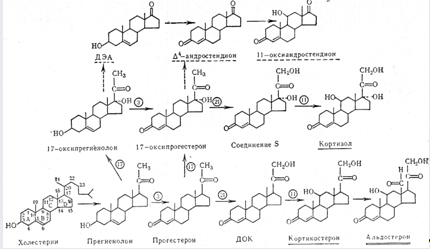

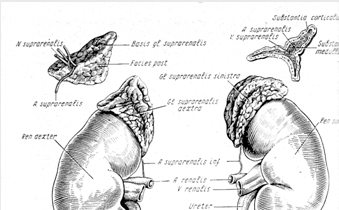

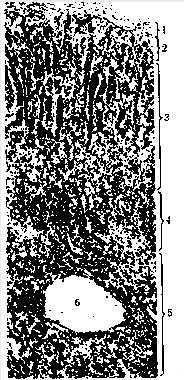
... контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою. Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо. Кіркова і ...
0 комментариев