Навигация
Волновой генетический код
177379
знаков
0
таблиц
29
изображений
СОДЕРЖАНИЕ
Актуальность темы
Пересмотр модели генетического кода
Расширение модели волнового генетического кодирования
Экспериментальные подтверждения существования волновых генов
Теоретические модели волновых генов
Математическое моделирование солитонов на ДНК
Экспериментальные доказательства солитонообразования на информационных биополимерах “in vitro”
Запись ИК-лазерного сигнала на уровне нелинейной динамики ДНК
О возможности создания лазера на информационных биомакромолекулах
Теоретическое исследование возможности создания лазера на Фрелиховских модах
Антенная модель, физико-математический формализм
Конверсия эпигеносигналов в электромагнитных солитонных структурах, их транспозиция в геном биосистем-акцепторов
Генератор пакетов уединенных волн (солитонов) в форме возврата Ферми-Паста-Улама
Единство фрактальной структуры ДНК-“текстов” и текстов на естественных языках
О возможности создания биокомпьютера на генетических структурах
Явление перехода света в радиоволны применительно к биосистемам и биокомпьютерам
Основные результаты
Литература
АКТУАЛЬНОСТЬ ТЕМЫ
В последние десятилетия начали постепенно выявляться некоторые кризисные явления в молекулярной биологии и биологии развития. После открытия структуры ДНК и детального рассмотрения участия этой молекулы в генетических процессах основная проблема феномена жизни - механизмы ее воспроизведения - осталась в своей сути не раскрытой. Отсюда ограниченность арсенала технических и биотехнических средств управления ростом и развитием биосистем. Наметился явный разрыв между микроструктурой генетического кода и макроструктурой биосистем. Даже открытие гомеобоксов ДНК, кардинально влияющих на формообразовательные акты эмбриогенеза, лишь более ярко высветили то, о чем в свое время предупреждал А. Г. Гурвич, считая, что нагрузка на гены слишком высока, и поэтому необходимо ввести понятие биологического поля, как пространственно-временной разметочной структуры, биологического поля, “... свойства которого... формально заимствованы... из физических представлений”. Таким элементарным полем, по Гурвичу, будет являться “... поле эквивалента хромосомы...”. И далее: “... хроматин сохраняет свою “активность”, т. е. является носителем активного поля, только в неравновесном состоянии”. Здесь видно предвидение лазерной накачки хромосом как типично неравновесного состояния, полученного нами in vitro спустя 50 лет для ДНК и нуклеогистона [18].
Близкие идеи мы видим и у А. А. Любищева в его работе 1925 г. “О природе наследственных факторов” . Он пишет: “Гены не являются ни живыми существами, ни кусками хромосомы, ни молекулами автокаталитических ферментов, ни радикалами, ни физической структурой, ни силой, вызываемой материальным носителем; мы должны признать ген как нематериальную субстанцию (выделено П.Г.), подобную эмбриональному полю Гурвича, но потенциальную” (выделено П.Г.). И далее: “... взаимодействие наследственности и хромосом подобно отношению материи и памяти по Бергсону... Гены в генотипе образуют не мозаику, а гармоническое единство, подобное хору” (выделено П.Г.). Через 3 года другой наш русский научный предшественник - В. Н. Беклемишев приходит к тем же идеям в своей работе, выполненной также в Перми, “Методология систематики”. Чтобы приблизиться к реальному морфопроцессу (эмбриогенезу), - необходимо принять идею музыки и речи как неких моделей векторов генетических актов. И в музыке, и в речи “... существуют “анатомические” свойства (возможны признаки стадий) - высота, интенсивность звука, обертоны и пр., а следовательно, возможно и описание отдельных стадий, и формальное описание процесса в его целостности... Музыкальная вещь аналогична морфопроцессу гораздо глубже, чем с первого взгляда кажется. Между обоими процессами бросается в глаза различие: изменения в развивающемся теле накапливаются, изменения в потоке музыки сменяются бесследно. Но истинным субъектом развития в музыке является эстетическое впечатление; оно растет и развивается под влиянием процесса звучания. Это морфопроцесс сложного духовного организма... Что является аналогом этого последнего в животных и растительных организмах? Не поток ли формативных раздражений, регулируемый индивидуальностью целого и направляющий морфогенез частей?”. Идеи русских биологов Гурвича, Любищева и Бекле-мишева - гигантское интеллектуальное достижение, намного опередившее свое время. Суть их мыслей в триаде:
1. Гены дуалистичны - они вещество и поле одновременно.
2. Полевые эквиваленты хромосом размечают пространство-время организма и тем самым управляют развитием биосистем.
3. Гены обладают эстетически-образной и речевой регуляторными функциями.
Современные молекулярная биология, генетика и эмбриология, проделав большой путь развития, завершили определенный виток в понимании сущности жизни. Оно было сугубо материалистичным, точнее, вещественным. Гены в этом смысле - только вещество. И когда это вещество - ДНК - детально изучили, открыв так называемый генетический код, то оказалось, что этого явно мало. Ключевая проблема биологии - преемственность поколений, наследственность, эмбриогенез - не раскрыта, более того, в тупике, правда более высокого ранга. Ситуация сейчас напоминает положение в классической физике начала xx века, когда с открытием элементарных частиц материи вещество вроде бы исчезло, осталось нечто, которое назвали неопределенным термином "энергия". Вот и в биологии, чем точнее понимание ДНК по части повсеместно принятой центральной догмы ДНК-РНК-Белок, тем дальше мы уходим от стратегии генома в построении биосистемы. Но если физика с достоинством приняла как реальность парадоксы: “здесь и там одновременно”, “волна и частица совмещены”, “электрон резонирует со всей Вселенной”, “вакуум - ничто, но он порождает все” и т. д., то биологии только предстоит пройти сходный путь (Дао Биологии), и он будет гораздо тяжелее. Фактически мы уже вышли на него, вовремя вспомнив мысли Гурвича, Любищева и Беклемишева.
Наша задача состояла и состоит в том, чтобы развить их концептуальную триаду в контексте современных знаний и полученных нами результатов по теории и практике разработки и использования механизмов и технических средств волновой коррекции биосистем.
Цель работы: показать возможность дуалистической трактовки работы генома эукариот на уровнях вещества и поля в рамках физико-математических моделей, соединяющих формализм явления солито-нообразования в ДНК на примере явления возврата Ферми-Паста-Улама, а также голографической и иной памяти хромосомного континуума как биокомпьютера, связанного с волей Творца.
Показать возможность обычных и “аномальных” режимов работы генома эукариот с использованием волновых образно-знаковых матриц, а также эндогенной и экзогенной (зависящей от Творца) семиотико-лингвистической компоненты.
Найти экспериментальные доказательства правильности пред-лагаемой теории волновых образных и образно-лингвистических матриц генома как структур стратегического управления метаболизмом высших биосистем.
Практическая направленность настоящего исследования:
в теоретико-экспериментальном обосновании феномена свертки, транспозиции и резонансного введения супергенетической информации от биодонора к биоакцептору; при этом передаваемые эпигеносигналы могут существовать как акусто-электромагнитные солитоны в рамках явления возврата Ферми-Паста-Улама и входить в семантические знаковые ряды генетических структур, также реализующиеся в форме солитонных возбуждений;
в теоретико-экспериментальном обосновании единства фрактальной структуры человеческой речи и текстовых структур генетических молекул ДНК и РНК; это положение заложило основу для разработки начальных основ методологии введения регуляторных квази-вербальных структур в виде модулированных электромагнитных солитонов непосредственно в геном растений;
в теоретико-экспериментальном обосновании возможности создания искусственных ДНК-логических устройств (биокомпьютеров) с исполь-зованием волновых (голографических и иных) принципов памяти, сравнимой по механизмам и возможностям с генетической;
в создании технических средств мягкого регуляторного вхождения в неизвестные ранее семиотические ареалы генома высших биосистем с целью лечения, создания гибридов, продления жизни людей, форми-рования организма человека как гармоничной и устойчивой к неблагоприятным факторам структуры.
Ранее нами предложена гипотеза эпигенетической кодовой иерархии уровней организации хромосомной ДНК, рибосом и внеклеточных матриксов высших биосистем и участия их в синтезе волновых образных фрактальных построений, используемых высшими биосистемами для собственной самоорганизации [25]. Нелинейная динамика (акустика) и связанные с ней электромагнитные излучения указанных биоструктур in vivo не случайны, взаимно коррелированы, носят биознаковый (в частности, речеподобный) характер, изоморфно отображают структурно-функциональные состояния каждой из обменивающихся волновыми сигналами организменных клеточно-тканевых подсистем. В пространстве-времени организмов в эпигенетическом режиме происходит обмен информацией по физическим каналам нелинейных акустическими электромагнитных колебаний. при этом стратегической компонентой рассматриваемых волновых знаковых рядов является акустическое и электромагнитное излучение совокупного генетического материала (генома) биосистем. В настоящей работе выдвинутые положения развиваются как трактовка волновых состояний (собственных физических полей) организма и попытка понимания биологического смысла явления генерации внутри и межклеточных полевых сигналов в качестве основы волновой и, вслед за этим, вещественной самоорганизации живых систем.
ПЕРЕСМОТР МОДЕЛИ ГЕНЕТИЧЕСКОГО КОДА
В настоящее время создалась парадоксальная ситуация с моделью генетического кода - вершиной достижений молекулярной биологии 60-х годов. Точность кодирования последовательностей аминокислот белков в этой модели странным образом уживается с двойной вырожденностью предлагаемого “кода” по линиям избытка транспортных РНК (тРНК) по сравнению с числом аминокислот и неоднозначного соответствия кодон-антикодон, когда только двум (а не трем) нуклеотидам триплетов иРНК необходимо точное спаривание c антикодоновой парой нуклеотидов тРНК, а по третьему нуклеотиду природой допускается неверное спаривание, так называемое “воблирование” (от англ. слова “wobble”- качание) по гипотезе Ф.Крика [4]. Это означает, что некоторые антикодоны могут “узнавать” более одного кодона в зависимости от того, какое основание находится в 1-м положении антикодона, соответствующем 3-му положению нуклеотида с учетом их антипараллельного комплементарного взаимодействия. “Узнавание” такого рода “неправильное”, если следовать парадигме генетического кода, поскольку возникают неканонические пары оснований “Аденин-Гуанин”, “Урацил-Цитозин” и другие с энергетически невыгодными водородными связями. “Код”, особенно митохондриальный, становится настолько вырожденным, и логически следующий отсюда произвол включения аминокислот в пептидную цепь столь велик, что как бы исчезает само понятие генетического кодирования.
Процитируем высказывание из книги Альбертса, Уотсона и др. “Молекулярная биология клетки” [20] (глава с характерным названием “Геном митохондрий имеет ряд поразительных особенностей”): “...в митохондриях обычные правила спаривания кодонов с антикодонами соблюдаются менее строго, и многие молекулы тРНК способны узнавать любой из четырех нуклеотидов в третьей (неоднозначной) позиции”. Вот эта “меньшая строгость”, как будто бы несовместимая с реально существующим метаболическим контролем порядка чередования амино-кислот в белках, заслуживает пристального внимания. “Меньшая строгость” не случайна, более того, она для чего-то нужна биосистемам.
Точность белкового синтеза эволюционно консервативна и высока, но может ли она достигаться такого рода “тайнописью”, когда “знак” (кодон) и “обозначаемое” (аминокислота) не всегда изоморфны, не однозначны? Если придерживаться старой догмы генетического кода, логично думать, что две разные аминокислоты, шифруемые двумя одинаковыми (третий не важен) нуклеотидами кодонов иРНК, будут с равной вероятностью включаться в пептидную цепь, т.е. случайно. И таких парных неоднозначностей даже в немитохондриальном коде насчитывается шесть, если не считать еще две по стоповым кодонам (они же “нонсенс” или бессмысленные). Так что же, существует “индульгенция разрешения” частых и случайных замен аминокислот при синтезе белков? Однако, известно, что такие случайные замены в большинстве случаев имеют самые отрицательные последствия для организма (серповидная анемия, талассемии и т.д.). Налицо явное противоречие: нужна точность (однозначность) отношений “знак-обозначаемое” (кодон-аминокислота), а придуманный людьми код ее не обеспечивает.
Поэтому существующее и общепринятое представление о ключевых (знаковых) механизмах синтеза белков нуждается в дополнительном анализе. В связи с этим более подробно рассмотрим предложенные в 60-х годах принципы генетического кодирования. Как оценили перечисленные и очевидные странности ведущие авторы теории и экспериментов в этой области - Ф.Крик, М.Ниренберг и их последователи? Основной узел противоречий - неоднозначные соответствия (кодон-аминокислота) приведены в таблице:
НЕОДНОЗНАЧНЫЕ СООТВЕТСТВИЯ И СИНОНИМО-ОМОНИМИЧЕСКАЯ ДВУМЕРНОСТЬ ГЕНЕТИЧЕСКОГО КОДА
![]()
Омонимичность
Видно, что пары разных аминокислот шифруются одинаковыми значимыми дублетами кодоновых нуклеотидов (“воблирующие” мало значимые, по Крику [4], и вообще нечитаемые, по Лагерквисту [11], нуклеотиды смещены в индекс). В терминах лингвистики это явление носит название омонимия, когда одни и те же слова имеют разный смысл (например, русские слова “лук”, “коса” или английские “box”, “ring” и т.п.). С другой стороны, избыточные различающиеся кодоны, обозна-чающие одни и те же аминокислоты, уже давно рассматривают как синонимичные.
В отношении омонимии генетического кода высказывания в литературе нам не известны. Таким образом, если считать дуплетно-триплетные кодоны “словами”, то сам код является, кроме прочего, двумерным, то есть омонимо-синонимичным. По этим измерениям код распадается, как это видно из таблицы, в основном, на парные семейства, избыточно, но не однозначно, шифрующие разные аминокислоты. И только в двух случаях из шести омонимичные дублеты обозначают близкие по структуре и функции аминокислоты (аспарагиновая-глутаминовая и аспарагин-лизин). Следовательно, при неоднозначном (ошибочном) выборе аминокислот высока вероятность синтеза аномальных белков, если следовать логике общепринятой модели кода. Большинство этих сомнений и наметок на будущее в мягкой форме уже прозвучало в обобщающей статье Ф.Крика и М.Ниренберга “Генетический код”. Процитируем авторов дословно ввиду стратегической важности обсуждаемых принципов генетического кодирования:
с.133: “белок ... является как бы длинным предложением, записанным с помощью двадцати букв”. Вот одно из первых и плодотворных сравнений белков, а затем и ДНК, с текстами естественных языков, сравнений, повсеместно принятых на первых порах лишь как метафора, а затем развитых и формализованных нами в качестве квази-речевых образований [14,25,26,29]. В этой замечательной аналогии зачаток будущего выхода из плоского и тупикового понимания природы генов, предтеча понятия образных кодов (слово как образ), а это согласуется с идеями Гурвича, Любищева и Беклемишева, которые также видели в хромосомах потенциальные волновые образные и даже эстетические структуры в качестве организующих биосистему начал. А.А.Любищев еще в 1925 году высказал предположение, что гены образуют не мозаику, а гармоническое единство, подобное хору [47]. Вслед за ним в 1928г. В.Н.Беклемишев [21] развил это, хотя и афористично, однако, с огромным предвидением, на десятилетия опередившим аналогию Крика и Ниренберга о белках как “предложениях”. Эмбриогенез он сравнил одновременно с музыкой и речью, в которых как и в дифференцирующихся тканях существуют “анатомические” свойства - признаки стадий: высота, интенсивность звука, обертоны и пр., и онтогенетические “эмбриологические” свойства - признаки хода процесса: ритм, мелодия и пр. Изменения в развивающемся теле накапливаются, а изменения в потоке музыки сменяются бесследно. Но истинным субъектом развития в музыке является эстетическое впечатление; оно растет и развивается под влиянием процесса звучания. Это морфопроцесс сложного духовного организма. Придя к этому, В.Н.Беклемишев спрашивает: что является аналогом этого последнего в животных и растительных организмах? Не поток ли формативных раздражений, регулируемый индивидуальностью целого и направляющий морфогенез частей?
Продолжим анализ основополагающей работы Крика и Ниренберга, постулирующей понятие генетического кода.
С.142 -143: “ ... до сих пор все опытные данные хорошо согласовывались с общим предположением о том, что информация считывается тройками оснований, начиная с одного конца гена. Однако, мы получили бы те же результаты, если бы информация считывалась группами в четыре или даже более оснований” или “...группами, содержащими кратное трем число оснований”. Это положение почти забыто или не понято, но именно здесь видно сомнение, обязательно ли код триплетный. И не менее важно, что предугадано будущее понимание текстов ДНК и РНК как смысловых фрактальных образований, родственных естественным языкам, что продемонстрировано в наших исследованиях [25,26,29].
С.153: “ ... одна аминокислота шифруется несколькими кодонами. Такой код называется вырожденным ... такого рода вырождение не говорит о какой-то неопределенности в построении молекулы белка ... оно лишь обозначает, что определенная аминокислота может быть направлена в соответствующее место цепи молекулы белка с помощью нескольких кодовых слов”.
Авторы видят, что синонимия еще не нарушает однозначности кода.
С.153 -154: Но дальше следует “...однако, все же имеется одна реальная возможность появления неопределенности при синтезе белка. Эта неопределенность могла бы возникнуть, если бы одно кодовое слово соответствовало нескольким аминокислотам. До настоящего времени был отмечен только один случай такой неопределенности. Белок, син-тезируемый поли-U, состоит не только из лейцина, но и из фенилаланина, причем на каждую молекулу лейцина приходится 20-30 молекул фенилаланина. При отсутствии в растворе фенилаланина поли-U использует лейцин в количестве, равном половине обычно используемого количества фенилаланина. Молекулярное объяснение этой неопределенности неизвестно”. Это первая и четкая констатация логического несовершенства предлагаемой модели кодирования, ее противоречия фактам. Затем, сомнения еще более усиливаются.
С.155: “некоторые кодовые слова почти наверняка состоят из трех оснований. Однако, 18 из 20 аминокислот могут быть закодированы словами, содержащими только два различных основания. Если же код все-таки троичный, то возможно, что в некоторых случаях правильное кодирование будет иметь место при условии, что из трех оснований считывается только два. Возможно, что такое несовершенство случается более часто в синтетических РНК-полимерах, содержащих одно или два основания, чем в естественных РНК-посредниках, которые всегда состоят из смеси всех четырех оснований. Поэтому результаты, полученные с помощью искусственных РНК, свидетельствуют лишь о кодовых возможностях клетки...”
Явно просматривается неуверенность, что код только триплетный, он может быть и дуплетным, и тетраплетным и даже гетеромуль-типлетным. Нам же представляется, в развитии этих сомнений, что кодовые возможности клетки, хромосом, ДНК не исчерпываются знаковыми тройками нуклеотидов. Как речеподобные структуры, нуклеиновые кислоты в составе хроматина способны к образованию in vivo метаязыков методом фрактализации, и поэтому кодирование белкового континуума может проходить через крупные блоки, шифрующие не только порядок включения отдельных аминокислот в пептид, но и последовательность создания белковых доменов, субъединиц и даже структурно-функциональных ансамблей ферментов, например, дыхательной цепи. Фрактальность в данном случае может пониматься и так: ДНК, РНК и белки - это разноязыкие тексты и то, что было в одном масштабе “фразой” или “предложением” в другом, более крупном, будет “словом”. Если еще укрупнять - “слово” превращается в “букву”. При более общем подходе можно рассматривать такие разномасштабные смысловые построения как знаки (иероглифы), являющиеся субстратом своего рода “информационного метаболизма” клеток. Такой путь образования метаязыков свойствен математике. У нас нет оснований думать, что геном не пользуется этим “математическим приемом” в полной мере, строя все новые усложняющиеся семиотико-семантические ареалы с их постоянными переобозначениями на разных уровнях организации биосистемы в процессе ее развития. При этом роль основной массы синтезирующихся в организме белков заключается в реализации метаболических конструкций, неявно закодированных в ДНК и имеющих квази-вербальную составляющую. Биосистему можно рассматривать как совокупность таких конструкций, и это находит определенное подтверждение в работах [25,26,29]. Такой ход рассуждений хорошо соответствует представлениям В.В.Налимова, считающего все живое частью Семантической Вселенной [49]. Человек, в соответствии с такой логикой, есть многообразие текстов, грамматику и семантику которых мы хотим охватить единым, вероятностно задаваемым взглядом. В.В.Налимов полагает, что личность является самочитаемым текстом - текстом, способным самоизменять себя. Уменьшая масштаб рассмотрения человека как самоорганизующейся системы и учитывая фрактальность (переходящую иногда в голографичность) его хромосомного континуума, можно считать, что обратное отображение человека в его собственный геном, как и отображения любого организма в его хромосомы, носит изоморфный текстово-образный характер [25,29]. Предлагаемый способ рассуждений призван показать прием логического выхода из ограничений первичной модели генетического кода, остановившейся в фазе слабого понимания правил орфографии "записи" белковых “слов” из аминокислотных "букв". Если же рассматривать идею фрактальности смысловых (текстовых) конструкций генома и принять их божественное начало, то подчеркнем, что эта идея восходит к VI в. и предложена Дионисием Ареопагитом в его труде “О божественных именах”. Он говорит, что печать Божественности (читай слова) лежит на каждом из нас, и “...оттиски Печати имеют много общего с ее оригиналом: оригинал присутствует в каждом из отпечатков весь, и ни в одном из них - лишь какой-то своей частью”. Частичность Печати определяется свойствами воспринимающего материала - конкретной личности, т.е. потенциально в каждого привносится все, идущее свыше, но расслышать, увидеть и понять это все целиком не под силу никому.
Неспособность ранней концепции генетического кода быть непротиворечивой, казалось, должна была побудить к поиску новых идей. Вместо этого предпочтение было отдано анализу механизмов точности белкового синтеза, но без главного мотива этой точности - механизмов выбора однозначностей из кодирующих дуплетов-омонимов. Вот образец этих, в данном аспекте бесполезных, описаний и рассуждений, но необходимых нам для иллюстрации псевдологики в оценке главного в генокоде [20]: “ ... точность белкового синтеза зависит от надежности двух адапторных механизмов: от связывания каждой аминокислоты с соответствующей молекулой тРНК и от спаривания кодонов в иРНК с антикодонами тРНК. Два механизма, действующие на этих этапах, совершенно различны. У многих аминоацил-тРНК-синтетаз имеется два отдельных активных центра: один ответственный за реакцию присоединения аминокислоты к тРНК, и другой, распознающий “неправильную” аминокислоту и удаляющий ее путем гидролиза. Точность спаривания кодона с антикодоном обеспечивается более тонким механизмом “кинетической коррекции”. После того как молекулы тРНК присоединят соответствующую аминокислоту, они образуют комплекс с особым белком, т.н. фактором элонгации (ФЭ,EF), который прочно связывается с аминоацильным концом молекулы тРНК и с молекулой GTP. Именно этот комплекс, а не свободная тРНК спаривается с надлежащим кодоном в молекуле иРНК. Связанный таким образом ФЭ обеспечивает возможность правильного спаривания антикодона с кодоном, но при этом препятствует включению данной аминокислоты в растущий пептид. Начальное узнавание кодона служит для ФЭ сигналом к гидролизу связанного с ним GTP до GDP+P, после чего ФЭ отделяется от рибосомы без тРНК и синтез белка продолжается. Благодаря ФЭ возникает короткий разрыв во времени между спариванием кодона с антикодоном и элонгацией пептида, что позволяет тРНК отделиться от рибосомы. “Неправильная” молекула тРНК образует в паре кодон - антикодон меньше водородных связей, чем правильная; поэтому она слабее удерживается на рибосоме и значит за данный промежуток времени имеет больше шансов отделиться”.
Комментируя эту, важную для нас, длинную выдержку, можно сказать, что акцент в ней сделан на взаимном узнавании тРНК и аминокислот через посредство аминоацил-тРНК-синтетаз. Механизм его не ясен. Что касается точности узнавания кодоном антикодона, то она иллюзорна в силу “воблирования” третьего нуклеотида, что уже обсуждалось. Представляется, что выбор из дуплетных кодонов-омонимов реализуется по резонансно-волновым и контекстным (ассоциативным, голографическим) и так называемым "фоновым механизмам" (см. ниже). До сих пор они находились вне экспериментов и рассуждений, но в настоящее время необходимость в этом очевидна. Омонимичность (неоднозначность) кода может быть преодолена точно так же, как это происходит в естественных языках ,- путем помещения омонима, как части, в целое, т.е. в законченную фразу, контекст которой дешифрует омоним и присваивает ему единственное значение, создавая однозначность. Поэтому иРНК в качестве своего рода “фразы” или “предложения” должна работать в белковом синтезе как функциональное кодирующее целое, задающее последовательность аминокислот на уровне ассоциатов аминоацилированных тРНК, которые комплементарно взаимодействуют со всей молекулой иРНК. При этом роль А,Р-участков рибосомы, если они реальны, заключается в акцепции таких ассоциатов - предшественников белка с последующей энзиматической сшивкой аминокислот в пептидную цепь. В этом случае будет происходить контекстно-ориентированный однозначный подбор бывших омонимичных дуплет-кодонов. Можно предсказать в связи с этим, что взаимодействие аминоацилированных-тРНК с иРНК носит коллективный фазовый характер по типу реассоциации (“отжига”) однотяжных ДНК при понижении температуры после “плавления” нативного полинуклеотида. Существуют ли экспериментальные данные, которые можно было бы трактовать в таком духе? Их немало и они сведены в обзорно-аналитическом исследовании [52]. Приведем некоторые из них.
Известно, что правильность узнавания молекулами тРНК терми-нирующих кодонов зависит от их контекстного окружения, в частности, от наличия за стоповым кодоном уридина и, кроме того, в работе убедительно показано следующее. Вставка строки из девяти редко используемых CUA-лейциновых кодонов после 13-го в составе 313 кодонов тестируемой мРНК сильно ингибируют их трансляцию без явного влияния на трансляцию других мРНК, содержащих CUA-кодоны. Напротив, строка из девяти часто используемых CUG-лейциновых кодонов в тех же позициях не имела выраженного эффекта на трансляцию. При этом ни редко, ни часто используемые кодоны не влияли на этот процесс, когда были введены после кодона 223 или 307. Дополнительные эксперименты продемонстрировали, что сильный позиционный эффект редко используемых кодонов не может быть объяснен различиями в стабильности иРНК или в степени строгости выбора соответствующих тРНК. Позиционный эффект становится понятным, считают авторы, если допустить, что транслируемые последовательности менее стабильны вблизи начала считывания: замедленность трансляции реализуется посредством малого использования кодонов, которые раньше следуют в сообщении, и это приводит к распаду продуктов трансляции, раньше чем осуществится полная трансляция. Как видим, для трактовки собственных экспериментов привлекаются громоздкие допущения о распаде продуктов трансляции, допущений, никак не следующих из их работы, и которые требуют специальных и тонких исследований. В этом смысле наша идея контекстных ориентаций в управлении синтезом белков проста, хотя экспериментально доказать ее непросто. Цитируемая работа хорошо высвечивает стратегическую линию влияния строго определенных и далеко расположенных от места образования пептидной связи кодоновых вставок в иРНК на включение или невключение конкретной аминокислоты в состав синтезируемого белка. Это именно дистантное влияние, но в цитируемой работе оно просто констатируется, оставаясь для исследователей непонятным и, видимо, поэтому даже не обсуждается. Таких работ становится все больше. В той, что мы обсуждаем, ссылаются, к примеру, на полдюжины аналогичных результатов, где трактовка в этом смысле также затруднена. Причиной этому является несовершенство общепринятой модели генетического кода. Это верно и потому, что имеются данные о существовании так называемого протяженного (swollen) антикодона [52]: во взаимодействии тРНК с иРНК в А-сайте рибосомы участвуют не три, а большее количество пар оснований. Это означает, что принятый пов-семестно постулат триплетности кода нарушается и здесь. Там же, в [52], приводятся результаты работы по взаимодействию тРНК-тРНК на рибосоме, и это соответствует нашей идее об ассоциате аминоацилированных тРНК как предшественнике белка. В [52] высказана мысль, что эффект действия контекста иРНК на однозначное включение аминокислот в пептид является отражением неких фундаментальных и пока плохо изученных закономерностей декодирования генетической информации в процессе белкового синтеза. В работе Ульфа Лагерквиста [11] “wobble”- гипотеза Крика получила расширенную трактовку и крайнее выражение, согласно которому нуклеотид в третьем положении кодона иРНК является лишним, бессмысленным, избыточным, его присутствие игнорируется, и поэтому чтение антикодоном кодона производится по правилу “два из трех”. Отсюда логично следует массированная неоднозначность прочтения иРНК и некорректность трансляции белковых молекул, что противоречит экспериментам, и это констатируется в [52], равно как и в других исследованиях. Вместе с тем, отмечается, что существует определенный уровень неоднозначности трансляции иРНК в клетке, но он слабо поддается осмыслению. Помимо ошибочной трансляции значащих кодонов и считывания стоп-кодонов как аминокислотных, в процессе белкового синтеза могут происходить многочисленные нормальные и редко ошибочные сдвиги и перекрытия рамок трансляции. Ошибки возникают в результате считывания дуплетов или квадриплетов оснований как кодонов. Механизмы сдвигов рамки считывания практически не изучены. Во многих работах показано, что ошибочная трансляции белков рибосомой вызывается разнообразными неблагоприятными факторами - антибиотиками, изменением температуры, созданием определенных концентраций катионов, аминокислотным голоданием и другими условиями внешней среды. Повышенная неоднозначность трансляции кодонов, локализованных в особом контексте, имеет биологическое значение и приводит к неслучайному распределению “ошибочных” аминокислот по длине синтезируемого полипептида, приводящему к модификациям функций белков с выходом на механизмы клеточных дифференцировок, и поэтому контексты иРНК являются субстратом естественного отбора. Оптимальный уровень “ошибок” трансляции (если это действительно ошибки) регулируется неизвестными механизмами, и он онтогенетически и эволюционно оправдан [52]. Этому соответствуют и наши экспериментально-теоретические данные [8-18] о волновых знаковых взаимодействиях в водно-жидкокристаллической среде клетки, в которые вовлечен белок-синтезирующий аппарат. Нами обнаружены резонансные частоты, общие для ДНК, рибосом и коллагена, и имеющие, вероятно, биознаковую природу, а также открыта способность хромосом и ДНК быть лазеро-активной средой [18].
Вернемся вновь к общепринятым поначалу основным положениям генетического кода: он является триплетным, неперекрывающимся, вырожденным, не имеет “запятых”, т.е. кодоны ничем не отделены друг от друга. И наконец, он универсален. Что осталось от этих положений? Фактически ничего. В самом деле, код, видимо, является двух-, трех-, четырех-, ... n-буквенным как фрактальное и гетеромультиплетное образование. Он перекрывающийся. Он имеет запятые, поскольку гетерокодоны могут быть отделены друг от друга последовательностями с иными функциями, в том числе с функциями пунктуации. Код не универсален - в митохондриях он приобретает специфические черты. Как понимать генетический код с учетом приведенных противоречий и предлагаемой нами логики рассуждений?
Для снятия этих противоречий можно постулировать качественную, упрощенную, первичную версию вещественно-волнового контроля за порядком выстраивания аминокислот в ассоциате аминоацилированных тРНК как предшественнике белка. С этой позиции легче понять работу генетического, а точнее белкового, кода как одной из множества иерар-хических программ вещественно-волновой самоорганизации биосистемы. В этом смысле такой код - первый этап хромосомных планов построения биосистемы, поскольку язык генома многомерен, плюралистичен и не исчерпывается задачей синтеза протеинов. Более детальное, физико-математически формализованное и экспериментально подтверждаемое, изложение новой версии работы белок-синтезирующего аппарата разра-батывается нами в настоящее время, хотя надо признать, что это задача xxI-xxII веков.
Основные положения предлагаемой ориентировочной модели вещественно-волновых знаковых процессов при биосинтезе белков сводятся к следующему:
1. Многокомпонентный рибонуклеопротеидный белоксинтезирую-щий аппарат является системой генерации высокоорганизованных знаковых семиотико-семантических излучений акустико-электромагнитных полей, стратегически регулирующих его самоорганизацию и порядок включения аминокислот в полипептидную цепь.
2. Аминоацилированные пулы тРНК ассоциируют в последователь-ности - предшественники синтезируемых белков до контакта с А-P участком рибосомы. При этом континуум антикодонов пула комп-лементарен всей иРНК, за исключением дислокаций, определяемых наличием неканонических нуклеотидных пар.
3. Порядок чередования аминоацилированных тРНК в ассоциатах-предшественниках белков определяется знаковыми коллективными резонансами всех участников синтеза аминокислотных последовательностей. Ключевые волновые матрицы здесь пре-иРНК, а также иРНК, работающие как целостный континуум разномасштабных по длине гетерополикодонов, включая интронную фракцию пре-иРНК как возможных макроконтекстов. Главная функция волновых матриц - ассоциативно-контекстная ориентация последовательности аминоацилированных тРНК, ориентация, в большей степени, чем воблгипотеза, игнорирующая правила канонических спариваний нуклеотидов в пространстве иРНК-тРНК.
4. На рибосоме, в дополнение и (или) наряду с резонансными регуляциями взаимного расположения кодон-антикодоновых континуумов функционируют лазероподобные излучения участников данного процесса, корригирующие порядок включения аминокислотных остатков в пептид.
5. Рибосома энзиматически ковалентно фиксирует “де-юрэ” пептидные связи аминокислотных последовательностей, намеченные “де-факто” в полиаминокислотном-поли-тРНК-ассоциате, как предшествен-нике белка.
6. Резонансно-волновая “цензура” порядка включения аминокислот в пептидную цепь устраняет потенциальный семантический произвол создания ошибочных белковых “предложений”, следующий из омонимии семейств кодонов, и обеспечивает их “аминокислотное осмысление” за счет контекстного снятия омонимии неоднозначных одинаковых дублетов в кодонах. Тот же механизм работает при неоднозначностях более высокого порядка, когда число кодонов (n+1).
7. Вырожденность генетического кода необходима для пре-иРНК-иРНК-зависимого контекстно-ориентированного точного подбора ацили-рованных тРНК, определяемого характером волновых ассоциативных резонансных взаимодействий в белок-синтезирующем аппарате.
8. Один из механизмов процесса создания безошибочных после-довательностей аминоацилированных тРНК на волновых матрицах пре-иРНК- иРНК можно рассматривать как частный случай частично комплементарной реассоциации однотяжных ДНК-ДНК и РНК-ДНК или, в более общем случае, как акт самосборки, известный для рибосом, хромосом, мембран и других молекулярно-надмолекулярных клеточных структур.
Таким образом, роль иРНК дуалистична. Эта молекула, как и ДНК, в эволюции знаменует собой узловое событие - взаимодополняющее синергичное расслоение вещественной и волновой геноинформации. Неоднозначность вещественного кодирования снимается прецезионностью волнового, которое реализуется, вероятно, по механизмам коллективных резонансов и лазерно-голографических (ассоциативных, контекстных) эффектов в клеточно-тканевом континууме [25,26,29]. Мега-контекстом здесь выступает словесно-волновое Божественное Начало. Скачок к более развитому волновому регулированию трансляции РНКБелок сопровождается частичным или полным отказом от правила канонического спаривания аденина с урацилом (тимином) и гуанина с цитозином, свойственного эволюционно ранее отобранным этапам репликации ДНК и транскрипции РНК. Такой отказ энергетически невыгоден в микромасштабе, однако информационно необходим, неизбежен и энергетически предпочтителен на уровне целостного организма.
Особо подчеркнем, что контекстные ассоциативно-голографические механизмы работы белок-синтезирующей системы организмов тесней-шим образом связаны с так называемым "Фоновым Принципом", который оказался универсальным и явился предметом крупного открытия [50]. С этой позиции макроконтексты пре-информационных и контексты информационных РНК можно рассматривать как фон, который обеспечивает резкое усиление сигнала, то есть выбора именно данной из двух омонимичных аминоацилированных тРНК, которая должна войти в белковую “фразу” или “слово”. Этот выбор возможен только после выделения когерентной составляющей в форме повторов одних и тех же осмыслений дублетов-омонимов в кодонах. Эту ситуацию можно пояснить на простом примере. Скажем, в предложении надо выбрать одно из двух слов (аналогов кодонов с дублетами-омонимами). Эти слова - “суд” и “сук”. Ясно, что выбор зависит от целого предложения, от контекста, который выступает как фон, позволяющий выделить сигнал - нужное слово. Если предложение звучит “я увидел толстый сук на дереве”, то замена здесь слова “сук” на “суд” будет равносильна введению шума и потере сигнала. Вероятно, аналогична роль пре-информационных РНК и интронов - это различные уровни контекстов, которые должны быть ка-ким-то образом “прочитаны” и “осмыслены” живой клеткой. “Субъектом чтения” может выступать многоликое семейство солитонов - оптических, акустических, конформационных, вращательно-колебательных и иных.
Функции таких солитонов могут выступать как способы регуляции кодон-антикодоновых знаковых взаимодействий. В качестве одного из способов можно представить солитонный механизм крутильных коле-баний нуклеотидов на сахаро-фосфатной оси иРНК, рассмотренный нами для однотяжных РНК-подобных участков ДНК [24]. Этот механизм “запоминает” последовательность нуклеотидов и может, вероятно, пере-давать информацию об этом дистантно, т.е. на расстояниях, существенно превышающих длину водородных связей. Без дальней (волновой) миг-рации сигнала о пре-иРНК-иРНК-последовательностях невозможна реализация ассоциативно-контекстных регуляций синтеза белков. Здесь необходима волновая континуальность, напрямую связанная с вкладом Божественного Начала как мегаконтекста, выступающего в форме естественного электромагнитного и акустического окружения земного шара. Первичная проверка предлагаемых положений может быть проведена относительно простым способом - по результатам влияния электромагнитных и акустических полей на синтез белков в бесклеточных рибосомальных системах, например с помощью ФПУ-генераторов и предполагаемых лазеров на ДНК [18,24,25,34,35].
Можно высказать предположение, что нарастающее увеличение людских так называемых внезапных смертей посреди видимого здо-ровья, приуроченных к зонам высоких уровней “электромагнитного СВЧ-смога”, зависит от нарушений тонкой волновой регуляции белко-вого синтеза. При этом могут образовываться аномальные белки “электромагнитного шока”, в том числе и ферментные системы синтеза эндерпинов ( эндогенных производных резерпина ), которые могут являться аномальными ко-факторами оксидоредуктаз, быстро блоки-рующими процессы внутриклеточной наработки энергии и, как следствие, летальный исход [28; неопубликованные результаты].
РАСШИРЕНИЕ МОДЕЛИ ВОЛНОВОГО ГЕНЕТИЧЕСКОГО КОДИРОВАНИЯ
В наших исследованиях [напр., 25] мы доказываем, что синтез бел-ка - лишь один из примеров генетического кодирования на волновом, стратегическом уровне, и трактуем генетическую память расширенно как солитонно-голографическую, свойственную геному-биокомпьютеру. Мы развиваем идеи Гурвича, Любищева и Беклемишева об излучениях хромосом, о “геноме-оркестре”. Действительно, если сравнить “запись” будущего организма на ДНК яйцеклетки с нотной записью, то одна музыкальная фраза способна ассоциативно восстановить всю совокупность музыкальных образов в нашей памяти, если мы хоть раз слышали мелодию. Принимая это, мы выходим на представления образного, знакового кодирования структуры организма последовательностями ДНК, т.е. они, последовательности нуклеотидов, являются чем-то вроде звучащих и видимых текстов, но не в поэтико-метафорическом смысле, а действительно текстами на неизвестных пока языках божественного происхождения в сложно-ритмической (музыкально-подобной?) волновой аранжировке. Но излучают ли хромосомы свет и звук? Эксперименты дают однозначно положительный ответ. Акустические поля хромосом, генерируемые как живыми клетками и их ядрами, так и выделенными из хромосом препаратами ДНК, сложно организованы, могут приобретать структуру солитонов, а главное, способны к дистантной трансляции гено-волновой информации [1,8,25,26-29]. Генетические молекулы дуалистичны - будучи веществом, они же работают как источники физических знаковых полей. Хромосомы, как главная знаковая фигура любой биосистемы, расщепляются на многомерные фрактальные семиотические структуры вещества и поля, закоди-рованные божественным промыслом.
Заметим, что зачатки этих идей и экспериментов возникли не на пустом месте (подробно об этом в [25] ). Первыми были, как упоминалось, Гурвич, Любищев и Беклемишев (20-е - 40-е годы), затем, через несколько десятилетий, в Новосибирске А.Н.Мосолов (1980г.), а затем группа ученых из Института общей физики АН (1984г.) с помощью световой и лазерной микроскопии обнаружили в клеточных ядрах (хромосомах) нейронов некие вибрирующие (звучащие) сферические образования. А.Н.Мосоловым было высказано предположение, что они являются источниками информационно-силовых генетических, а точнее, эмбриональных полей в духе идей А.Г.Гурвича, но с существенной поправкой: во-первых, это не фотонные поля, во-вторых, обнаруженные звуковые излучения, по Мосолову, имеют голографическое проис-хождение. Это была первая четко сформулированная гипотеза знако-несущих (образных) волновых голографических структур генома высших биосистем. Эту гипотезу мы развили на основании собственных исследований.
Мы шли несколько иным путем, пытаясь на первых этапах доказать правильность физико-математической модели Инглендера, предложенной им в 1980г., об особых волновых состояниях ДНК-солитонах. Затем теоретических моделей солитонов появилось множество, но никто до 1991г. солитонов на ДНК экспериментально не обнаружил. В 1985г. методом спектроскопии корреляции фотонов нам удалось зафиксировать необычные аномально долго затухающие колебания (звук) ДНК in vitro с меняющимся спектральным составом, особым образом распределенным во времени. Это наблюдение было настолько необычно, что было принято за экспериментальную ошибку и поэтому забыто на 6 лет до тех пор, когда мы вновь повторили эту работу. Было обнаружено, что ДНК обладает способностью как бы в автоматическом режиме (квази-спонтанно) синтезировать “незамолкающую сложную мелодию с повторяющимися музыкальными фразами” [8,25,29]. Такие повторы по ряду признаков походили на солитонный процесс в форме явления так называемого возврата Ферми-Паста-Улама (ФПУ), а сами колебания ДНК сродни тем, что наблюдали Мосолов и др.
Коротко о возврате ФПУ. Если в цепочке осцилляторов (маятников), соединенных пружинками с нелинейными связями, возбудить один из них, то возникнет необычное колебание с повторениями (возвратами) энергии первоначального возбуждения. Это своего рода “память” всех нелинейных систем, свойственная и молекулам ДНК, что продемонстрировано на уровне теоретической модели А.А.Березиным. Но в ДНК такая память, как показали наши исследования, приобретает особое значение. Она может нести семиотическую нагрузку и выступать в форме своего рода “волновых генов” - солитонов с внутренней колебательной структурой, сходной, вероятно, с голограммами [25]. Однако, для избирательного “чтения” in vivo генетических голограмм на уровне солитонов и в пределах жидкокристаллического хромосомного континуума биосистемы необходимо лазерное поле хромосомного аппарата. Долгие годы его пытались найти и воспроизвести вне живой клетки. В принципе, нам это удалось. Мы получили лазерное излучение на препаратах ДНК и хромосом методом двухфотонно-возбуждаемой люминесценции [14].
ЭКСПЕРИМЕНТАЛЬНЫЕ ПОДТВЕРЖДЕНИЯ СУЩЕСТВОВАНИЯ ВОЛНОВЫХ ГЕНОВ
Теперь о возможностях волнового (эпигенетического, суперге-нетического) уровня работы хромосом и его реализации через технические устройства. В 1957г. в Китае исследователь Дзян Каньджэн начал, а с 70-х г.г. в России продолжил супергенетические эксперименты, которые перекликались с предвидениями Гурвича, Любищева и Беклеми-шева. С 60-х г.г. в Новосибирске академиком В.П.Казначеевым и его школой начаты исследования, призванные подтвердить идеи дистантных волновых знаковых межклеточных взаимодействий. Ими был открыт так называемый зеркальный цитопатический эффект, когда культуры живых клеток и тканей, герметично разделенных кварцевым стеклом, обмениваются волновой регуляторной информацией, связанной с функциями генетического аппарата.
Реальные и достоверные эксперименты в области волновой генетики первым начал проводить Дзян Каньджэн. Итоговые работы его известны. Прибор Дзян Каньджэна, дистантно (десятки сантиметров) передающий “волновые гены” от донора к реципиенту, использует собственные излу-чения биосистем-доноров, причем, как считает автор, только в СВЧ-дипазоне электромагнитных полей. Авторское теоретическое обоснова-ние эффектов, полученных с помощью этой аппаратуры, нуждается в существенной доработке. Однако, экспериментальные данные убеди-тельны. Это “волновые гибриды” пшеницы и кукурузы, земляного ореха и подсолнуха, огурца и дыни, утки и курицы, козы и кролика. Полученные гибридами признаки передаются по наследству. Блестящий эмпирик Дзян Каньджэн не объясняет тонкие механизмы открытых им эффектов, но это нисколько не умаляет значимость результатов, суть которых в доказательстве реальности волновых генов.
ТЕОРЕТИЧЕСКИЕ МОДЕЛИ ВОЛНОВЫХ ГЕНОВ
Возникла настоятельная необходимость в теоретическом развитии идеи волнового генома, в физико-математическом и теоретико-биоло-гическом осмыслении работы хромосом и ДНК в полевом и вещественном аспектах. Первые попытки решить эту задачу предприняли П.П.Гаряев и А.А.Березин (Отдел теоретических проблем РАН), а также А.А.Васильев (Физический институт РАН). В основу их представлений были положены принципы когерентных физических излучений, голографии и солитоники, теория физического вакуума, фрактальные отображения структур ДНК и человеческой речи. Суть идей Гаряева - Березина - Васильева “ГБВ-модель” [25,19,30,33,53] состоит в том, что геном высших оранизмов рассматривается как солитонный биоголографический компьютер, формирующий пространственно-временную структуру развивающихся эмбрионов по каскадам реестров волновых образов-предшественников. При этом в качестве носителей полевых генов выступает континуум волновых фронтов, задаваемый мультиплексными геноголограммами, образуемыми гелевым жидкокристаллическим хромосомным континуумом. Акт “считывания” информации осуществляют сами же хромосомы, генерирующие лазерные свет и звук в широких диапазонах. Близкую роль играют также и солитоны на ДНК - особый вид акустических и электромагнитных полей, продуцируемых генетическим аппаратом самого организма и способных к посредническим функциям по обмену стратегической регуляторной информацией между клетками, тканями и органами биосистемы. Важно также и то, что квази-голографические решетки, в том числе входящие в состав колебательных структур солитонов, являются лишь частным простейшим случаем кодово-образной информации, зафиксированной в хромосомном континууме организма. Мультиплетнокодовая ДНК, где аминокислотный код - только малая часть, и “эгоистическая ДНК” хромосом анализируются в рамках ГБВ-модели как потенциальный стратегический информационный вектор всех клеток и тканей организма, включая кору головного мозга. Геном работает не только на вещественном, но и на волновом, на “идеальном” (тонкоматериальном) уровне. Эта идеальная компонента, которую можно назвать супергено-континуумом, и является главной знаковой фигурой генома, обеспечивающей развитие и жизнь человека, животных, растений, а также их программируемое естественное умирание. Вместе с тем важно понять, что нет резкой и непреодолимой границы между волновыми и материальными уровнями хромосом. Оба они образуются вещественными матрицами, но гены дают материальные реплики в виде РНК и белков, а супергены преобразуют падающие на них эндо- и экзогенные физические поля, формируя из них пространственно-временные разметочные волновые структуры. Более того, гены могут быть составной частью голографических решеток супергенов и регулировать их полевую активность. И наконец, супергены могут формироваться как ДНК-РНК-нуклеопротеид-лазерное поле, промодулированное их текстами.
Особого внимания заслуживает в ГБВ-модели обоснование единства фрактальной (повторяющей самою себя в разных масштабах) структуры последовательностей ДНК и человеческой речи. То, что четыре буквы генетического алфавита (Аденин, Гуанин, Цитозин, Тимин) в ДНК-текстах образуют геометрические фрактальные структуры, конста-тировано Джефри в 1990г. в рамках так называемого “хаотически-игрового” математического представления последовательностей нуклеотидов. Это не вызвало особой реакции научной общественности. Однако, обнаружение нами геноподобных фрактальных (в геометрическом аспекте) структур в человеческой речи, и не только в буквенных рядах русских и английских текстов, но и в последовательностях слов этих текстов методически нетривиально. Хотя сама идея фрактальности естественных текстов не нова, но это идея смысловой фрактальности. В целом, такой ход мысли созвучен направлению в семиотике, называемому “лингвистическая генетика”, направлению, которое пытается объяснить некоторые, похоже общие закономерности создания гибридов биосистем и “гибридов” слов. Становится понятнее принятое, и уже привычное, опережающее сравнение ДНК с естественными текстами, имевшее преимущественно метафорический характер. Мы разработали модель фрактального представления естественных (человеческих) и генетических языков [14], которая позволяет предположительно считать, что “квазиречь” ДНК обладает потенциальным неисчерпаемым запасом “слов”. В этом пункте мы существенно расходимся с известными представлениями об исключительно трехбуквенном шифровании молекулой ДНК и только последовательностей аминокислот в белках. Предлагаемая фрактальная модель может стать полезной для тонкого количественного и смыслового сравнения знаковой структуры любых текстов, в том числе генетических. Можно попытаться подойти к дешифровке семантических построений ДНК и, соответственно, к составлению алгоритмов речевого или квазиречевого обращения к геному любой биосистемы через аппаратуру, моделирующую знаково-волновые функции генетического аппарата. Первичная практическая проверка ГБВ-модели в области “речевых” характеристик ДНК дала положительные результаты. Так же, как и в экспериментах Дзян Каньджэна, был получен эффект прямой трансляции и введения геноволновой информации от донора к акцептору. Затем мы создали устройства - генераторы солитонных полей Ферми-Паста-Улама (ФПУ), в которые можно было вводить речевые алгоритмы, к примеру, на русском и английском языках. Такие вербальные структуры превращались в особые электромагнитные (солитонные) модулированные поля - аналоги тех, которыми оперируют клетки в процессе волновых коммуникаций. Организм и его генетический аппарат в определенных, не совсем понятных, условиях “узнает” такие “волновые фразы” как свои собственные и в ряде случаев поступает в соответствии с введенными человеком извне речевыми управляющими воздействиями. Не исключен также фактор экзобиологического контроля за работой геноструктур через аналогичные коммуникативные каналы. А если быть точным, то этот контроль есть Божественное Начало. Видимо, геном не само-достаточен для управления организацией биосистемы.
Нам удалось получить предварительные результаты по влиянию кодовых вербальных структур, транслируемых через аппаратуру, на геном растений-акцепторов. зафиксирован факт распознавания геномами растений человеческой речи, что коррелирует с идеей лингвистической генетики о глубинном сходстве механизмов словообразования и синтеза речи для хромосом и человеческих языков [48], соответствует гипотезе существования праязыка людей [44] и перекликается с постулатом структурной лингвистики, по которому все естественные языки имеют глубинную врожденную универсальную грамматику, инвариантную для всех языков [51]. И, вероятно, для языка генома как одного из них. Об этом же говорят широко известные данные о генетическом дефиците хромосом, не позволяющем полностью реализовать программы развития организма в условиях внешней искусственной полевой информационной изоляции. Фильтрация или искажение некоторых (гено-знаковых) внешних естественных физических полей вызывает уродства и гибель эмбрионов. Это означает, что коммуникации генетических субстратов с экзогенными волновыми знаковыми структурами безусловно необходимы для гармоничного развития организма. Внешние Божественные (или искусственные) волновые сигналы несут дополнительную, а может быть и главную, информацию в геноконтинуум Земли. Такая идея в какой-то мере подтверждается нашими прямыми экспериментами, которые показали, что ДНК в состоянии жидкого кристалла может являться неким подобием антенны для приема сигналов явно искусственного характера, резко отличного от штатных акустических излучений ДНК. Этот факт, возможно фундаментального характера, проявляется в том, что молекулы ДНК в режиме “приема”, длящегося не один час, начинают вести себя аномальным образом, имитируя квази-разумное поведение на уровне нелинейной динамики полимера, что регистрируется методом корреляционной лазерной спектроскопии и прямым наблюдением за броуновским движением моле-кул [10,15,25]. Не исключено, что в этом выявляются высшие регуляторные волновые супергеносигналы, предназначенные для стратегического уп-равления организмами Земли.
Рассмотрение генетических структур как космических волновых антенн хорошо согласуется с идеями Хозе Аргуэльеса относительно генетического кода. Он считает, что последний описывает лишь часть общей картины жизни, и дополнением к нему является свет - лучистая энергия. Это резонансная лучистая инфраструктура - световое тело - входит в диапазон излучения, который управляется кодом Цолькина, гармонического модуля майя. Отслеживая “источник” лучистой энергии, Аргуэльес приходит к мнению, что он является ядром нашей Галактики. Излучаемые им спиральные потоки пульсаций вращаются в прямом и обратном направлениях и представляют собой код, контролирующий самопередающее и самопреобразующее свойства лучистой энергии. Описываемый гармоническим модулем майя галактический код является первоисточником, пропитывающим и наполняющим жизнью код ДНК.
Совсем недавно нам удалось получить еще одно свидетельство в пользу существования волновых генов. Был открыт феномен генерации широкополосного спектра радиополей в диапазоне от килогерц до одного мегагерца молекулами ДНК in vitro в особых условиях. [42].Препараты ДНК возбуждались в специальной резонансной системе He-Ne лазера со специ-фической модуляцией светового пучка (длина волны - 632,8 нм). Есть основания полагать, что такая система превращения видимой области спектра электромагнитного излучения в радиодиапазон свойственна биосистемам и они используют эти сверхслабые радиоизлучения в качестве носителя волновой (квазиречевой) генетической информации. Феномен генерации радиоволн из красного света оказался универсальным, свойственным и неорганическим веществам, и сейчас он многократно перепроверяется. Однако, уже сейчас зафиксирован особый спектральный состав радиоволн, генерируемых с участием генетических структур. Надо полагать, что обнаружен один из волновых “языков” генома, где субъектами “чтения” и “озвучивания” генотекстов выступают солитонные волны (бризеры) в ДНК [25,40,42].
Еще одно подтверждение нашей трактовки кодовых функций генома получено в 1994г. американскими исследователями [12]. Работая с “кодирующими” и “некодирующими” последовательностями ДНК эука-риот (в рамках старых представлений о генах), эта научная группа из Бостона пришла к выводу, противоречащему догме о том, что знаковые функции сосредоточены только в белок-шифрующих участках ДНК. Они применили метод статистического анализа естественных и музыкальных текстов, известный как закон Ципфа-Мандельброта, и принцип избы-точности текстовой информации Шеннона, рассчитываемый как энтропия текстов (относительно энтропии текстов и статистики распределения слов в текстах см., например, [17]). В результате они получили, что “некодирующие” районы ДНК более схожи с естественными языками, чем “кодирующие”, и что, возможно, “некодирующие” последователь-ности генетических молекул являются основой для одного (или более) биологических языков. Кроме того, авторами был разработан статистический алгоритм поиска кодирующих последовательностей ДНК, который выявил, что белок-кодирующие участки обладают существенно меньшими дальнодействующими корреляциями по сравнению с зонами, разделяющими эти участки. Распределение ДНК-последовательностей оказалось настолько сложным, что использованные методы переставали удовлетворительно работать уже на длинах, превышающих 103 - 102 пар оснований. Распределение Ципфа-Мандельброта для частот встреча-емости “слов” с числом нуклеотидов от 3 до 8 показало большее соответствие естественному языку некодирующих последовательностей по сравнению с кодирующими. Еще раз подчеркнем, что кодирование авторы понимают как запись информации об аминокислотной последовательности, и только. И в этом парадокс, заставивший их заявить, что некодирующие регионы ДНК - это не просто “junk” (в переводе с английского - “мусор”), а структуры, предназначенные для каких-то целей с неясным пока назначением. Дальнодействующие корреляции в этих структурах авторам также непонятны, хотя и обнаружена нарастающая сложность некодирующих последовательностей в эволюции биосистем, что продемонстрировано на примере семейства генов тяжелой цепи миозина при переходе от эволюционно низких таксонов к высоким. Эти данные полностью соответствует нашим идеям о том, что именно “некодирующие” последовательности ДНК, т.е. около 95 - 98 % генома, и являются стратегическим информационным содержанием хромосом. Оно имеет материально-волновую природу и поэтому многомерно и, по своей сути, выступает как ассоциативно-образная лингвистиковолновая программа эмбриологического начала, смыслового продолжения и логического конца любой биосистемы. Поняв это, авторы с ностальгической грустью прощаются со старой и хорошо послужившей моделью генетического кода, не предлагая, правда, ничего взамен.
Еще одна фундаментальная особенность голографии, экстраполи-рованная на биосистему, дает большую ясность в понимании волновых механизмов “самоанализа” биосистемы. Так, открытый Денисюком “принцип относительности в голографии” (доплеровская голография) выявил способность интерферограмм, записывающих движущиеся в трехмерном пространстве объекты, как бы предсказывать их пространственное положение в будущем. Если доплеровская голограмма формируется волной, отраженной от движущегося объекта, то обращенная такой голограммой волна, идя обратным ходом, фокусируется не на сам объект, а несколько впереди его. При этом существенно, что точка фокусировки обращенной волны является в этом случае именно той точкой, в которую переместится объект за время, пока обращенная волна распространится от голограммы до этого объекта. Нет оснований считать, что принцип относительности в голографии не применим к биосистеме, если сама голография уже используется организмом в мозговой памяти. Этот принцип может являться элементом оценки динамики метаболических процессов и “слежения” за движущимися внутриклеточными структурами и за крупномасштабной динамикой морфогенетических тканевых перестроек. Доплеровская система эндогенного биоконтроля дает способ элементарной прекогниции метаболических событий. С этим перекликается другое, близкое описываемым, свойство голограмм. Доказано, что с голограмм возможно считывание сигнальных импульсов с обращенной временной и пространственной структурой и продемонстрировано, что порфириновые компоненты таких важнейших биомолекул как гемоглобин и хлорофилл в полистирольной матрице могут голографически записывать разнесенные во времени лазерные импульсы. При считывании воспроизводится как относительная задержка, так и временная форма записанного сигнала. Таким образом, в принципиальном плане можно представить уже не только внутреннее динамическое пространственное “самоотсле-живание” биосистемой самой себя, но и аналогичный контроль за структурой собственного времени с анализом коротких временных отрезков, направленных как в прошлое, так и в будущее.
Работы по обращению временного сигнала голограммой важны и как пример, что средой памяти такого рода могут служить ключевые биомолекулы живых систем. И это не случайно. Фотосинтез (хлорофилл) и дыхание (гемоглобин) - первоистоки жизни на Земле, а структура времени для биосистем также важна для них как структура собственного пространства, и контроль за ними может осуществлять фундаментальный волновой принцип интерференции и дифракции.
Порфирины - не единственный бионоситель голографической памя-ти. Аналогично работает сложный фоточувствительный белок микробных клеток бактериородопсин. Следующим важнейшим бионосителем голо-графической информации является производное коллагена - желатина. Этот субстрат с 1968 года стал классическим объектом для изучения механизмов формирования амплитудных и амплитудно-фазовых голо-графических решеток в различных диапазонах электромагнитных полей. Использование производных коллагена подтверждает обсуждавшуюся выше мысль о том, что система внеклеточных матриксов, структурнофункциональной основой которых является коллаген, работает с использованием собственной памяти на интерферирующие поля и (или) способна к синтезу эпигенознаковых дифракционных решеток типа псевдоголограмм без участия интерферирующих полей.
Не исключено, что в клетках и тканях используется тепловой диапазон эндогенных полей для автосканирования и записи. Известно, что для записи на желатине используется ИК-СО![]() лазер (длина волны 10,6 мкм), который вызывает в ней локальные необратимые конфор-мационные переходы типа спираль-клубок, связанные со структурными состояниями гидратационной воды. Существенным свидетельством правильности голографической парадигмы, кроме наших исследований, служат работы Будаговского и Евсеевой, показавших в прямых экспериментах возможность дистантной трансляции биологически активного морфогенетического голографического сигнала с растения-донора на каллусную ткань растения-акцептора близкого вида .
лазер (длина волны 10,6 мкм), который вызывает в ней локальные необратимые конфор-мационные переходы типа спираль-клубок, связанные со структурными состояниями гидратационной воды. Существенным свидетельством правильности голографической парадигмы, кроме наших исследований, служат работы Будаговского и Евсеевой, показавших в прямых экспериментах возможность дистантной трансляции биологически активного морфогенетического голографического сигнала с растения-донора на каллусную ткань растения-акцептора близкого вида .
Возможно, неким приближением к тому, что происходит в биосистеме и коррелирует с упоминавшимися работами, служат также исследования, в которых обнаружено, что гели коллагена обладают способностью каномально долгому затуханию собственных макроконформа-ционных колебаний, давая при этом повторяющиеся, но разноплановые фурье-спектры, что нами подтверждено и развито в теоретическом и экспериментальном планах не только для коллагена, но и для ДНК и рибосом. Этот феномен, возможно, связан с солитонообразованием на информационных биополимерах в форме явления возврата Ферми-Паста-Улама. Свойство аномально малой затухаемости колебаний коллагена находит довольно неожиданное подтверждение в электроакустике костей. Обнаружена спонтанная генерация переменных электрических волн костной тканью даже тогда, когда она взята у мертвых животных, спустя многие часы после их смерти. Заманчиво объяснить это явление колебаниями коллагеновых фибрилл в составе костной ткани и генерацией ими полей за счет своих электретных свойств, известных для коллагена. Если это правильно, получает объяснение еще один необычный факт: пленки-подложки из коллагена, используемые как искусственный внеклеточный матрикс при выращивании на них культуры фибробластов, при укалывании иглой начинают организовывать упорядоченные движения фибробластов. Последние собираются в четкие ритмические паттерны, причины возникновения которых не ясны. И здесь можно проследить явление того же порядка, что и в случае генерирующей поля изолированной костной ткани. В обоих случаях имеют место квазиспонтанные колебания гелей коллагена, порождающие акустические и электрические поля, которые дополнительно возбуждаются уколом во втором случае. Система коллаген-фибробласты in vitro будет в таком случае элементарной моделью матрично-клеточных морфогенетических отношений, когда запускаются механизмы клеточно-тканевых движений по программам волновых фронтов акустико-электромагнитных голограмм динамичной системы “клетки U внеклеточный матрикс” с жидкокристаллическими компонентами, способными помнить интерферирующие поля.
Теоретически информационная емкость голографических решеток даже в двумерном варианте при записи электромагнитных колебаний огромна, так как они несут восьмимерную информацию. Объем голог-рафической памяти в биосистеме (помимо мозга) может быть еще большим за счет записи в трехмерной жидкокристаллической среде так называемых мультиплексных голограмм, когда меняются отношения интенсивности опорного и объектного пучков и меняются углы между ними, что логично предположить в мобильной тканевой среде организма.
Расшифровка механизмов быстрой и безинерционной передачи больших массивов волновой информации в организме позволяет по иному взглянуть на проблемы онкологии. Действительно, трудно иначе объяснить известные эксперименты по индукции опухолей имплантированными в ткань шлифованными (отражающими волны) инородными материалами. Шероховатые инородные предметы вызывают опухоли в 12% случаев по сравнению с 49% зеркальных того же состава. В этом случае переродившиеся клетки, дающие клоны опухолевых, возникают в соединительно-тканной капсуле, окружающей инородное тело, или редко за пределами капсулы, но они никогда не обнаруживаются в монослое клеток, лежащих непосредственно на инородном теле. Для естественных эндогенных электромагнитных и акустических полей организма, отражающие их инородные тела являются шумовыми помехами в передаче волновой информации по голографическим и солитонным механизмам.
Как один из путей нового понимания генома нами было начато изучение некоторых трудно интерпретируемых феноменов жизненных форм. К числу таких необычных и непонятных (“аномальных”) явлений относятся эффекты следовой памяти генетического материала, обнаруженные нами и независимо группой Роберта Пекоры (США). Сюда же относится феномен так называемого фантомного листового эффекта, подтвержденного во многих лабораториях, в том числе и нами. Такую память генома можно рассматривать как один из видов генетической полевой памяти биосистем на молекулярно-ткане-органном уровне. Она реализуется одновременно как ассоциативно-голографическая и как память последействия ДНК и дает иные версии работы хромосом, дополняющие уже известные механизмы и переводящие проблему биологического морфогенеза в иные гносеологические планы. Эта проблема нами исследована одновременно в теоретико-биологическом, физико-мате-матическом и экспериментальном планах [8,25,27,37]. В связи с этим представляется, что существует геносемиотический сектор работы хромосомного континуума, в котором происходит дуалистическое расщепление смысловых рядов ДНК на уровни вещества (реплики РНК и белков, знаковые топологии хромосом) и поля (знаковые акустика и электромагнитные излучения генома). Исходя из этого, кодирующую иерархию хромосомного аппарата эукариот можно представить следующим образом.
Вещество: хромосомная ДНК как кодирующая структура, в которой триплетный генетический код выполняет первичные простейшие гено-знаковые функции синтеза иРНК и белков (1-й уровень). Хромосомная ДНК, включающая спейсерные и интронные зоны как многомерная структура знаковых фрактальных топологических форм жидкого крис-талла, частным случаем которых выступают голографические решетки полиядерного когерентного континуума генома (2-й уровень). Квази-“речевые” фракталы полинуклеотидных ДНК-РНК-последовательностей, более длинных чем триплеты кодонов и белковых генов и кодирующих на “словесно”-образном уровне (3-й, 4-й... n-й уровни).
Поле: отчужденные от генома в форме волновых знаковых построений “идеальные” или “смысловые” (образные) ряды, субъектом генерации и “понимания” которых выступает геном как солитонноголо-графический биокомпьютер с квази-”речевыми” атрибутами, и соответственно, квазисознанием. Назначение волновых и “речевых” команд заключается в логической квази-сознательной разметке потенциальной биосистемы, т. е. в синтезе ее полевого относительно устойчивого и вместе с тем динамичного “автопортрета” - волновой физической матрицы для правильного распределения вещества организма в его собственном пространствевремени. В этом плане логично рассмотреть:
а) информационные отношения между системой внеклеточных матриксов, цитоскелетом, белок-синтезирующим аппаратом и хромосомами с новых позиций, учитывающих собственные экспериментальные данные об изоморфных волновых состояниях этих биоструктур;
б) вклад эндогенных физических полей в биоморфогенез;
в) роль эндогенных физических полей в эмбриогенезе биосистем с точки зрения солитоники и голографии; высказана идея изоморфно-гомоморфных отображений на уровне полевых функций генома с его способностью к солитонным возбуждениям и транспорту их по “водному” клеточно-межклеточному континууму.
В рамках проведенных математических экспериментов обнару-жилась способность компьютерных математических моделей солитонов Инглендера-Салерно-Маслова на ДНК запоминать последовательности нуклеотидов, отображая их в динамике собственного поведения во времени. При этом обозначилась и очевидная обратная задача - если солитоны осуществляют “запоминание” структур ДНК в своих амплитудно-траекторных модуляциях, то естественно считать практически возможной генерацию этой информации за пределы ДНК, что коррелирует с нашими экспериментами и теорией по дистантной передаче волновых морфогенетических сигналов [25]. В физическом и семиотическом планах это может и должно найти отображение в форме ретрансляции солитонами последовательностей нуклеотидов (на уровне крупных блоков) в адекватной читаемой, в том числе и человеком, форме.
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ СОЛИТОНОВ НА ДНК
Марио Салерно первым начал компьютерное экспериментирование с солитонами на ДНК не только как с формальными математическими структурами, он попытался связать их поведение в одномерном прост-ранстве полинуклеотидов с их биогенетическими, а точнее, с эпиге-нетическими функциями. При этом он развил первую модель солитонов на ДНК, предложенную Инглендером и соавторами. Эта модель и в последующем ее более детальные формы, включая нашу (см. ниже), представлена в понятиях механических систем как цепочка осцилляторов (оснований ДНК), связанных упругими нелинейными сахаро-фосфатными связями. Вслед за Салерно основное внимание мы уделили реально существующим известным последовательностям ДНК и влиянию их на характер поведения солитонов. На первом этапе мы повторили его эксперименты, но на существенно более длинных отрезках ДНК. Действительно, солитонные возбуждения типа кинков чувствительны к месту своей инициации, и продвижение их вдоль одной из цепочек ДНК, когда они раскрыты вследствие тепловых флуктуаций, сопровождается специфической модуляцией траектории кинков во времени. Такие солитоны являются структурами, излучающими электромагнитное и акустическое поле, их внутренняя колебательная структура способна отобразить и ретранслировать тексты и иные знаковые структуры ДНК во внутри- и внеклеточное пространство, по крайней мере на уровне крупных блоков последовательностей. В качестве примера можно привести поведение кинка на фрагменте ДНК длиной 1020 пар оснований из вируса саркомы птиц.
C-район ДНК (1
![]() 1020 нуклеотид) на 3’-конце вируса саркомы птиц. Содержит несколько “семантически” определенных участков, таких, как полипептид-кодирующий участок (между 558 и 675 нуклеотидами); PolA (936) - 3’-конец вирусной РНК, сайт поли-аденилирования; 916 нуклеотид - 5’-конец вирусной РНК (“capping site”); Red-участок (917 - 936) - короткий концевой повтор вирусного генома; Pro - вероятный компонент промотора транскрипции (между 870 и 900); палиндром-”шпилька” (870 - 912).
1020 нуклеотид) на 3’-конце вируса саркомы птиц. Содержит несколько “семантически” определенных участков, таких, как полипептид-кодирующий участок (между 558 и 675 нуклеотидами); PolA (936) - 3’-конец вирусной РНК, сайт поли-аденилирования; 916 нуклеотид - 5’-конец вирусной РНК (“capping site”); Red-участок (917 - 936) - короткий концевой повтор вирусного генома; Pro - вероятный компонент промотора транскрипции (между 870 и 900); палиндром-”шпилька” (870 - 912).
На рис.1 и рис. 2 кинки имеют форму пиков “горных гряд”, а не ступенек, поскольку взята производная от функции уравнения синус-Гордона. Здесь горизонтальная ось - последовательность ДНК, верти-кальная - амплитуда солитона. Ось на зрителя - время. Видно, как при изменении места инициации солитона на определенных последо-вательностях полинуклеотида заметно меняется динамика этой уеди-ненной волны в форме ее колебательных движений вдоль цепочки ДНК.
Исследуемый район молекулы богат функционально (и семантически) биологически значимыми участками, и мы вправе ожидать, что они, эти участки, будут изменять, модулировать, то есть вводить ДНК “текстовую” информацию в солитонную волну как в переносчик генетических сообщений. Такая модуляция колебательной структуры солитонов отчетливо наблюдается на приведенных графиках. Можно полагать, что спектральный состав частот колебаний солитонов является одним из механизмов преобразования текстовых структур ДНК и РНК в волновую форму и средством передачи генетических и иных сообщений в одномерном пространстве вдоль цепочек полинуклеотидов и (или) в трехмерном измерении генома как отдельной клетки, так и тканевого континуума биосистемы.
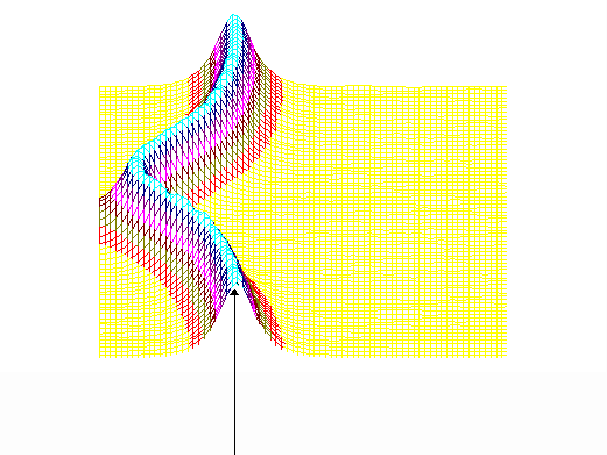
400-ый
Рис.1
Влияние нуклеотидной последовательности ДНК на динамику конфор-мационного возмущения уединенной (солитоноподобной ) волны. Последо-вательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 400-ый нуклеотид.
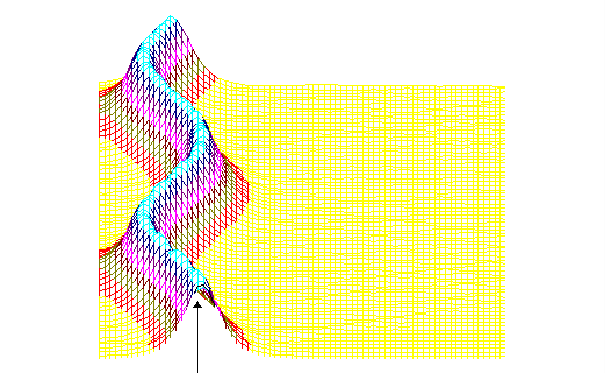
450-ый
Рис.2
То же, что на рис.1, но центр возмущения цепочки ДНК на 450-ом нуклеотиде.
Так работает компьютерная модель динамики солитонов, в определенной мере развитая Салерно после ее выдвижения Инглендером. Салерно дал формализм, описывающий вращательные колебания нуклеотидов молекулы ДНК, для того чтобы объяснить экспериментальные данные по водородно-тритиевому обмену в ДНК. Согласно этой модели по Инглендеру, в цепи ДНК могут возникать (под воздействием теплового шума) и распространяться открытые состояния (“плавление” двойной спи-рали ДНК на коротких участках, обогащенных АТ-парами ) в виде локализованных дислокаций ( уединенных волн). Марио Салерно, про- должая работу Инглендера, в упрощенном варианте выявил влияние последовательности нуклеотидов на нелинейную динамику вращательных колебаний нуклеотидов на однотяжных участках ДНК, образующих такие открытые ("open state") области. Позднее Якушевич, Федянин, Хомма и др. рассмотрели различные обобщения модели Инглендера, с оценкой особенностей строения ДНК, учитывая обрыв водородной связи при открытии оснований, парность цепи ДНК и другие степени свободы, отличные от вращательных. Однако, в указанных работах недостаточно сказано о причинах возникновения дислокаций в ДНК. Мы предлагаем возможный механизм этого процесса в ДНК, альтернативный гипотезе Инглендера о воздействии теплового шума как причины раскрытия пар оснований. Мы считаем, что дислокации на ДНК могут возникать при изменении периода спирали ДНК (основная часть идеи принадлежит М.Ю.Маслову).
В нашей модели нуклеотиды ДНК рассматриваются как осцилляторы, подвешенные на невесомом нерастяжимом стержне; сахаро-фосфатная связь между соседними нуклеотидами в цепи моделируется линейными пружинами; спирализация вдоль цепи не учитывается; водородные связи между комплементарными основаниями моделируется “гравитационным” потенциалом. Гамильтониан по М. Салерно выглядит следующим образом: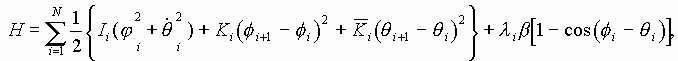 (1)
(1)
где: ![]() - углы вращений нуклеотидов в разных цепях,
- углы вращений нуклеотидов в разных цепях, ![]() - константы упругости вдоль цепей,
- константы упругости вдоль цепей,
![]() - число пар в цепи,
- число пар в цепи,
![]() - момент инерции оснований,
- момент инерции оснований,
![]() - константа упругости водородных связей между комплементарными основаниями.
- константа упругости водородных связей между комплементарными основаниями.
Коэффициенты ![]() в уравнении (1) определяются в соответствии с правилом:
в уравнении (1) определяются в соответствии с правилом:
![]() в случае АТ и ТА пар,
в случае АТ и ТА пар,
![]() в случае ГЦ и ЦГ пар;
в случае ГЦ и ЦГ пар;
![]() - параметр, определенный Федяниным и Якушевич и полученный на основе модели синус-Гордона и экспериментальных данных. Далее для упрощения модели считается, что
- параметр, определенный Федяниным и Якушевич и полученный на основе модели синус-Гордона и экспериментальных данных. Далее для упрощения модели считается, что
![]()
Уравнения движения для разности
![]() , полученные из (1), имеют по М. Салерно вид:
, полученные из (1), имеют по М. Салерно вид:
![]() (2)
(2)
где произведена замена
![]() .
.
В случае
![]() , в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона:
, в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона:
![]() , (3)
, (3)
”непрерывный аналог” системы (2). Это уравнение имеет солитонные решения, в частности, односолитонное решение, или кинк, соответствует дислокации в цепи.
Основным предположением моделей Инглендера-Салерно является то, что взаимодействие между комплементарными основаниями описывается потенциалом ![]() (4), в котором не учитывается обрыв водородной связи.
(4), в котором не учитывается обрыв водородной связи.
В нашей работе рассматривается следующий потенциал :
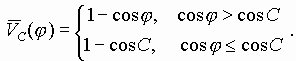
Кроме того, учитывается вязкость водной среды (в воде вязкость
![]() ~ 1).
~ 1).
Рассматриваются также факторы, приводящие к спирализации ДНК, при этом они считаются внешними силами, задаваемыми потенциалом
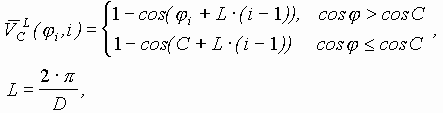
где ![]() - период спирали.
- период спирали.
Уравнения (2) с потенциалом ![]() и с учетом вязкости принимают вид:
и с учетом вязкости принимают вид:
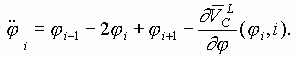 (5)
(5)
Известно, что период спирали ДНК меняется в зависимости от влажности. В частности, для кристаллической ДНК
![]() , а в водной среде
, а в водной среде ![]() - в пределах от 10. 3 до 10. 6. Именно этим фактором обусловлено явление суперспирализации. При изменении шага спирали в цепи ДНК (с фиксированными или замкнутыми концами) возникает напряжение, связанное с недостатком (избытком) количества витков спирали до релаксированного состояния. Если
- в пределах от 10. 3 до 10. 6. Именно этим фактором обусловлено явление суперспирализации. При изменении шага спирали в цепи ДНК (с фиксированными или замкнутыми концами) возникает напряжение, связанное с недостатком (избытком) количества витков спирали до релаксированного состояния. Если ![]() , то при переходе из сухого в увлажненное состояние для цепи длиной в 300 пар оснований возникнет избыток в
, то при переходе из сухого в увлажненное состояние для цепи длиной в 300 пар оснований возникнет избыток в
![]() витка.
витка.
В нашей работе на основе результатов численного моделирования, представленных ниже, выдвигается следующая гипотеза: изменение шага спирали может привести не только к суперспирализации, но и к локальному распариванию цепи ДНК. Кроме того, при суперспирализации напряжение в цепи снимается не полностью, поэтому локальное распаривание, вероятно, может происходить и одновременно с суперспирализацией.
Система (5) численно интегрировалась в интервале ![]() с шагом
с шагом
![]() . Начальные условия следующие:
. Начальные условия следующие:
![]()
![]()
![]()
Период спирали в системе (5)![]() длина poly(A)-цепи - 300 пар оснований. То есть параметры периода спирали в начальных условиях и в системе (5) различны. Таким образом смоделирован перенос ДНК из кристаллического состояния в увлажненное.
длина poly(A)-цепи - 300 пар оснований. То есть параметры периода спирали в начальных условиях и в системе (5) различны. Таким образом смоделирован перенос ДНК из кристаллического состояния в увлажненное.
Граничные условия следующие (назовем их “квазициклическими”):
![]()
Особенностью данной модели является то, что при переходе из состояния с периодом в 10 пар в состояние с периодом в 10, 5 пар почти вся цепь оказывается денатурированной (“расплавленной”). Приведенные ниже результаты описывают процесс ренатурации такой цепи с возникновением дислокаций.
В этих экспериментах варьировались параметры: 1) диссипация ![]() 2) отношение параметров упругости
2) отношение параметров упругости
![]() 3) угол обрыва водородных связей
3) угол обрыва водородных связей
![]() .
.
На рис. 3 и 4 представлены результаты численного интегрирования системы (5). Показана не сама функция ![]() , а разница
, а разница
![]() , поскольку область изменения функции
, поскольку область изменения функции
![]() (приблизительно от
(приблизительно от
![]() до
до ![]() ) велика по сравнению с характерными изменениями в системе (приблизительно от 0 до 9). Горизонтальная часть графиков соответствует нераспаренному участку цепи с периодом спирали
) велика по сравнению с характерными изменениями в системе (приблизительно от 0 до 9). Горизонтальная часть графиков соответствует нераспаренному участку цепи с периодом спирали
![]() . Наклонная часть графиков на рис. 3(a), 4(а) соответствует дислокации.
. Наклонная часть графиков на рис. 3(a), 4(а) соответствует дислокации.
Можно сделать следующие выводы:
1) Способность к образованию дислокации в этой модели сильно зависит от
![]() . При
. При
![]() дислокация возникла во всех рассмот-ренных случаях.
дислокация возникла во всех рассмот-ренных случаях.
2) Способность к образованию дислокации также сильно зависит от параметра![]() . Во всех случаях, когда параметр
. Во всех случаях, когда параметр ![]() велик (
велик (![]()
на рис. 1.а, 2.а ), дислокация возникла. В пользу этого утверждения также свидетельствует сравнение рис. 3(а) и 4(г).
Как показывают дополнительные расчеты, влияние![]() на эффект проявляется в меньшей степени. Дислокация образуется или не образуется вне зависимости от значения
на эффект проявляется в меньшей степени. Дислокация образуется или не образуется вне зависимости от значения ![]() (
(![]() или
или
![]() ). При больших значениях
). При больших значениях ![]() дислокация образуется медленнее, чем при меньших.
дислокация образуется медленнее, чем при меньших.
3) На рис. 3(а), 4(в,г) видно, что дислокация имеет кинкообразную форму.
Ширина дислокации зависит от параметров ![]() (чем больше
(чем больше
![]() , тем меньше ширина дислокации) и
, тем меньше ширина дислокации) и
![]() (чем больше
(чем больше
![]() , тем меньше ширина дислокации).
, тем меньше ширина дислокации).
Развивая дальше модели солитонных возбуждений в ДНК (совместно с М.Ю.Масловым и др.) мы использовали условия, при которых цепочки ДНК моделируются набором ровибронных осцилляторов, подвешенных на невесомом нерастяжимом стержне; для простоты спирализация цепи не учитывается, а ровибронные степени свободы одной из цепочек считаются “замороженными”.
В этом случае гамильтониан для “активной” цепочки записывается в следующем виде:
H=H0+H1+H2
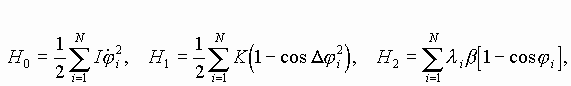 (1)
(1)
где: ![]() - число пар оснований в цепи;
- число пар оснований в цепи;
![]() - гамильтониан, описывающий собственные осцилляции мономеров (
- гамильтониан, описывающий собственные осцилляции мономеров (![]() - углы вращения нуклеотидов в цепочке,
- углы вращения нуклеотидов в цепочке,![]() - момент инерции оснований);
- момент инерции оснований);
![]() - гамильтониан , характеризующий нелинейно-периодическую связь между осцилляторами (
- гамильтониан , характеризующий нелинейно-периодическую связь между осцилляторами (![]() - константа упругости цепочки,
- константа упругости цепочки,
![]() ),
), ![]() - гамильтониан,
- гамильтониан,
 (а)
(а)
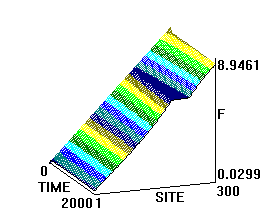 (б)
(б)
а)x0=200 б)x0=250
Рис.3
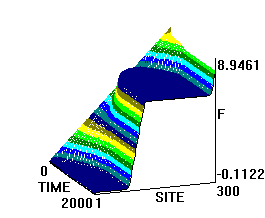 в)
в) 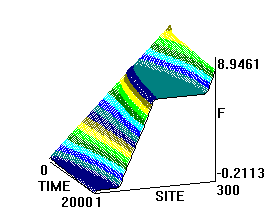 г)
г)
в) x0=300 г) x0=350
Рис. 4
описывающий нелинейную связь между “активной” и “замороженной” (![]() ) цепочками ДНК (
) цепочками ДНК (![]() - константа упругости водородных связей между комплементарными основаниями, коэффициенты
- константа упругости водородных связей между комплементарными основаниями, коэффициенты ![]() в уравнении (1) определяются в соответствии с правилом:
в уравнении (1) определяются в соответствии с правилом:
![]() в случае АТ и ТА пар,
в случае АТ и ТА пар,
![]() в случае ГЦ и ЦГ пар;
в случае ГЦ и ЦГ пар;
![]() - параметр, полученный ранее (см. выше) и определяемый на основе модели синус-Гордона).
- параметр, полученный ранее (см. выше) и определяемый на основе модели синус-Гордона).
При малых
![]() гамильтониан
гамильтониан![]() , что совпадает с соответствующей частью общего гамильтониана, использованного ранее (см. выше). В этом случае уравнения движения для
, что совпадает с соответствующей частью общего гамильтониана, использованного ранее (см. выше). В этом случае уравнения движения для ![]() , полученные из (1),
, полученные из (1),
имеют вид:
![]() (2)
(2)
где произведена замена
![]() .
.
В случае
![]() в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона:
в системе (2) можно перейти к безразмерному дифференциальному уравнению синус-Гордона:
![]() , (3)
, (3)
”непрерывный аналог” системы (2). Это уравнение имеет солитонные решения, в частности, односолитонное решение, или кинк, характеризующий динамику распространения дислокации в цепи.
В соответствии с (1) система нелинейных уравнений движения записывается следующим образом:
![]() (4)
(4)
Как видим, системы (2) и (4) существенно различаются. Отметим, однако, что проведенное нами численное моделирование динамики систем (2) и (4) показало следующее: если в качестве начальных условий для численного интегрирования (2) выбрать односолитонное решение его “непрерывного аналога” (3) - кинк (см. выше), то обнаруживается принципиальное сходство в характере решений.
Однако, при задании начальных условий в следующем виде:
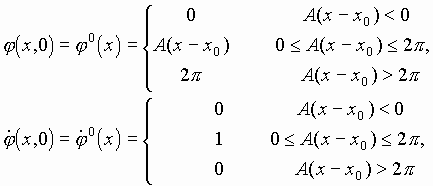 (5)
(5)
где ![]() - ”ступенчатая” функция с высотой ступени
- ”ступенчатая” функция с высотой ступени
![]() и углом наклона уступа A, выявилось различие динамики данных систем (срав. рис.1 и 2,3). Более точно, системы (2) и (4) численно интегрировались методом Рунге-Кутта четвертого порядка с начальными условиями, заданными в виде (7), в интервале
и углом наклона уступа A, выявилось различие динамики данных систем (срав. рис.1 и 2,3). Более точно, системы (2) и (4) численно интегрировались методом Рунге-Кутта четвертого порядка с начальными условиями, заданными в виде (7), в интервале ![]() с шагом
с шагом
![]() . Граничные условия - “квази-циклические”:
. Граничные условия - “квази-циклические”:
![]()
![]() (поли-A-последовательность). Параметр системы
(поли-A-последовательность). Параметр системы ![]() . Варьировался параметр A (угол наклона уступа функции
. Варьировался параметр A (угол наклона уступа функции
![]() ).
).
Численное интегрирование системы (2) ( рис. 1) показало, что образуются две уединенных волны, движущихся справа налево по цепи с постоянной скоростью. Первая волна имеет форму квазикинка, а вторая волна имеет форму квазибризера, причем скорость первой волны превосходит таковую для второй. Обе волны за счет “квазициклических” граничных условий, доходя до левого конца, появляются на правом конце без изменения своей формы. Квазикинк, проходя по цепи маятников, изменяет координату каждого маятника на угол
![]() (маятник делает полный оборот). Поэтому, проходя по замкнутой цепи маятников К раз, он изменяет координату каждого маятника на угол
(маятник делает полный оборот). Поэтому, проходя по замкнутой цепи маятников К раз, он изменяет координату каждого маятника на угол
![]() Этим объясняется “уступообразная” форма графика на рис. 1.
Этим объясняется “уступообразная” форма графика на рис. 1.
На рис. 2 представлены результаты интегрирования системы (4) при тех же условиях. Из рисунка видно, что образуются те же две уединенных волны - квазикинк и квазибризер. Но принципиальное отличие от рассмотренного случая состоит в том, что квазикинк в самом начале движется с отрицательным ускорением, так что в результате его скорость оказывается меньше скорости квазибризера. Заметим, что исследования проводились на однородной поли-A-последовательности; так что изменение скорости квазикинка нельзя объяснить влиянием неоднородности цепочки. Этот эффект объясняется нелинейным взаимодействием между ее мономерами.
Рис. 3 иллюстрирует результаты интегрирования системы (4) при тех же условиях за исключением того, что A=2. В данном случае реализуется только квазикинк и его отрицательное ускорение в начале движения таково, что в результате он движется в направлении, противоположном первоначальному. При интегрировании системы (2) в аналогичных условиях также образуется только квазикинк. Его скорость не меняется по сравнению со случаем рис. 1.
Существенно, что при соответствующих условиях в системе типа ДНК или РНК могут возникнуть перевзбужденные ровибронные состояния. На квантовом языке это было бы адекватно перезаселению высоко лежащих квантовых уровней по сравнению с основным (реализации инверсной заселенности). В этом случае возникает заманчивая мысль, связанная с принципиальной возможностью создания биосолитонного лазера (БСЛ) на молекулах ДНК.
Однако, в теории динамики биополимеров хорошо известно, что конформационные движения реализуются по механизму ограниченной диффузии ввиду сильного влияния диссипативных сил со стороны микроокружения. По этой причине решение проблемы создания БСЛ на ДНК представляется весьма проблематичным, по крайней мере, для подтверждения идеи необходимо выполнение условий:
![]() где
где ![]() и
и ![]() - ширина и скорость солитона соответственно,
- ширина и скорость солитона соответственно, ![]() - время диссипации. Положив
- время диссипации. Положив
![]()
![]() 5 A и
5 A и
![]() (скорость звука), имеем оценку
(скорость звука), имеем оценку
![]() . Отметим, что характерное время диссипации за счёт водных гидродинамических сил
. Отметим, что характерное время диссипации за счёт водных гидродинамических сил
![]() а время затухания, обусловливаемое процессами внутри самой молекулы
а время затухания, обусловливаемое процессами внутри самой молекулы
![]() (см., напр., Шайтан К.В. Биофизика. М.,1994. Т.39. С.949.; Чернавский и др. 1986. № 287. С. 21.).
(см., напр., Шайтан К.В. Биофизика. М.,1994. Т.39. С.949.; Чернавский и др. 1986. № 287. С. 21.).
Существует также и другая сложность в отношении самосогласования биосолитонов и волны электромагнитного переизлучения. Напомним, что математическое моделирование в данном случае проводилось на монотонной поли-A ДНК и поэтому оставалось неясным влияет ли гетерогенная естественная последовательность ДНК на динамику солитонного возбуждения в молекуле. Чтобы проверить это, как и ранее, был взят С-район ДНК на 3’-конце вируса саркомы птиц в качестве полигона для запуска солитонов на разных участках полимера. На этот раз вычисляли производную от функции с тем, чтобы нагляднее показать движения солитонов.
На рис.5,6 (см. ниже) хорошо видно, как при сдвиге области возбуждения солитонной волны от правой нижней части графика налево траектория волны претерпевает существенные изменения, т.е. “словесно-речевое” наполнение ДНК отображается в поведении солитона. Но главное здесь не только и не столько в этом. На этот раз характерно не качание волны около некоторого положения равновесия, а движение ее в левую часть цепочки после определенного временного интервала. В этом видится определенный биологический смысл. Солитон как потенциальный “субъект чтения” ДНК должен “просматривать” протяженные контекстные зоны, а не застревать на одних и тех же “словах”.
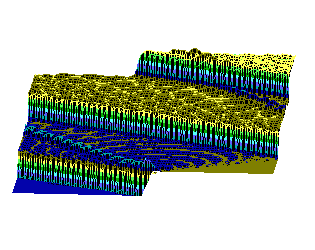 а)
а)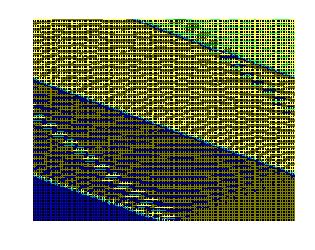 б)
б)
Рис.5
а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (2) при значении параметра A=1.
б) То же, вид сверху.
 а)
а)
 б)
б)
Рис.6
а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (4) при значении параметра A=1.
б) То же, вид сверху.
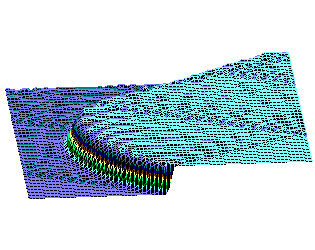 a)
a)
 б)
б)
Рис.7
а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (4) при значении параметра A=2.
б) То же, вид сверху.
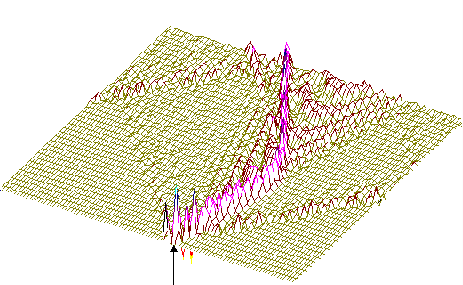
200-ый
Рис.8
Солитонное возбуждение ДНК, но с учётом нелинейности ковалентных связей в сахаро-фосфатном остове ДНК. Последовательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 200-ый нуклеотид.
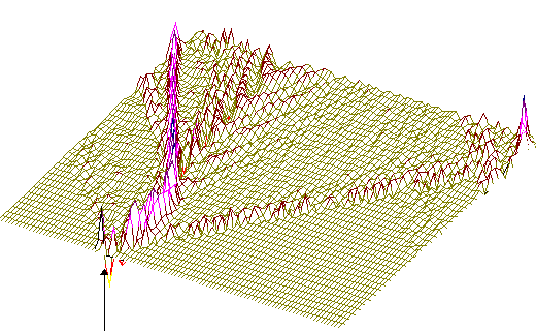
400-ый
Рис.9
То же, что на рис. 8, но центр возмущения - 400-ый нуклеотид.
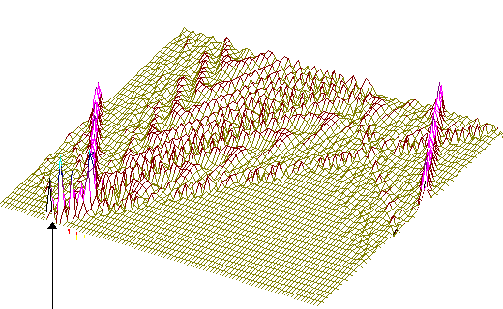
500-ый
Рис.10
То же, что на рис. 9, но центр возмущения - 500-ый нуклеотид.
ЭКСПЕРИМЕНТАЛЬНЫЕ ДОКАЗАТЕЛЬСТВА СОЛИТОНООБРАЗОВАНИЯ НА ИНФОРМАЦИОННЫХ БИОПОЛИМЕРАХ “IN VITRO”
Способны ли молекулы ДНК и белков к солитонным возбуждениям, предсказанным в многочисленных теоретических моделях? Нами предприняты попытки фиксации нелинейных волн такого рода in vitro методом спектроскопии корреляции фотонов. Выявлены устойчивые эффекты, которые по ряду признаков соответствуют, в частности, процессу спонтанного солитонообразования в рамках явления возврата Ферми-Паста-Улама. [8,19,25,31,32]. Обнаружилось, что при переходе от разбавленного раствора ДНК к полуразбавленному можно зарегистрировать аномально долго затухающие акустические колебания гелевого континуума ДНК. Слабо затухающие колебания исчезают по мере перехода от полуразбавленного к разбавленному раствору и в результате уменьшения длины фрагментов ДНК. Эти данные подтверждают ранние работы для агарозы и коллагена, где впервые обнаружен феномен аномально слабой затухаемости плотностных колебаний биогелей. Аномальное поведение ДНК зарегистрировали после наших наблюдений и японские авторы методом прямой регистрации броуновской динамики флуоресцентно-меченой ДНК. Причем, в работе японцев выявились и другие необычные особенности нелинейной динамики ДНК, которые не укладываются в хорошо разработанные теоретические модели Цимма и Роуза, но которые хорошо соответствуют нашим наблюдениям и трактовке молекул ДНК как структур, резонирующих на особые внешние волновые регуляторные сигналы [25,6,7,15,16,29] (также см. ниже). Такая самоорганизация волновых процессов в ДНК может происходить и при таких физических условиях, когда существенную роль играют кооперативные процессы на уровне макромолекулярного континуума молекул ДНК, приближающегося к структуре хромосом. Чем более структура растворов ДНК отличается от архитектоники ДНК в хромосомах (в приводимых нами экспериментах это относительно короткие фрагменты полимера), тем менее существенны коллективные дальние (в масштабах макромолекулярных протяженностей полинуклеотида) взаимодействия между цепями ДНК, столь важные для эпигенетических функций генома. Ключевым звеном в данных экспериментах является четкая регистрация поведения ДНК in vitro, которое ранее зафиксировано Бреннером и Носсалом для агарозы и коллагена в аналогичных условиях. Это позволяет рассматривать нелинейную динамику такого рода для ДНК и других информационных биополимеров как проявление солитонных свойств в рамках явления возврата Ферми-Паста-Улама (ФПУ). Нелинейная динамика ДНК, ее гидродинамическое поведение и акустика чрезвычайно чувствительны к внешним физическим воздействиям in vitro - энзиматической рестрикции, разбавлению-концентрированию, нагреву-охлаждению, ультразвуковой обработке, слабым механическим воздействиям, облучению ИК-лазерным полем, излучением ФПУ-генератора с широкополосным электромагнитным спектром. Эти и аналогичные воздействия могут и должны в той или иной мере оказывать влияние на генетический аппарат в условиях in vivo, искажающее нормальные эпигенознаковые функции хромосом, что также подтверждается в наших экспериментах. Нелинейная динамика ДНК обнаруживает и другие “аномальные” свойства. Мы зафиксировали резкое различие коэффициентов диффузии для кольцевых и линеаризованных плазмидных ДНК [33], которое также не укладывается в циммовскую теорию поведения полимеров в водных растворах и в этом плане находит подтверждение в работах группы Роберта Пекоры (США) и упоминавшемся исследовании Матсумото с соавторами. Эти необычные свойства ДНК, вероятно, играют важную роль, например, для понимания механизмов управляемого “пилотирования” и точной “посадки” транспозонов ДНК (аналогов плазмид) в пределах жидкокристаллического сверхвязкого и сверхплотного континуума хромосом. Эта задача находится в области общей и нерешенной проблемы молекулярной биологии - проблемы самоорганизации внутриклеточных, межклеточных и межтканевых структур, их “взаимоузнаваний”. Ясно, что, зная волновые, гидродинамические и иные механизмы точного пилотирования таких немаловажных для человека транспозонов, как онкогены и обратнотранскриптазный геном вируса иммунодефицита человека, мы будем иметь возможность корректировать их в необходимом направлении, исключающем патогенез. Не менее существенным представляется факт обнаружения нелинейной динамики ДНК с признаками поведения солитонов по типу явления возврата ФПУ. Это также дает вклад в осознание принципов макромолекулярных и надмолекулярных взаимоузнаваний в организме по линии солитонно-резонансных дальних взаимодействий и делает более реалистичной попытку дать новую версию работы генома эукариот, обсуждавшуюся выше. Мы обнаружили и другие необычные проявления физических свойств ДНК - ее последействие или следовую память [25]. Этот феномен ставит проблему новых типов геномных функций. Возможно, это явление тесно связано с особой памятью генома высших биосистем, а также, вероятно, и с памятью коры головного мозга. Но если для ассоциативной корковой памяти и памяти генома растений нами и другими даны физико-математические модели в терминах и понятиях голографических и солитонных процессов, то память последействия ДНК - явление далеко не ясное и нуждающееся в более глубоком исследовании и осторожной трактовке. Этот эффект зарегистрирован нами при динамическом лазерном светорассеянии на препаратах высокоочищенных ядер из эритроцитов кур и на высокополимерной чистой ДНК из зобной железы теленка [25].По сути, аналогичное явление наблюдала группа Р.Пекоры (США) и назвала его “MED-effect” (Mimicing Effect of Dust), т. е. эффект, имитирующий пыль. Так же как и в наших работах, этот феномен обнаружен методом корреляционной лазерной спектроскопии на рестриктных фрагментах ДНК строго определенной длины. И в этом случае ДНК вела себя “аномальным”образом: зондирующие фотоны дифрагировали не только на полинуклеотидных цепях, но и на “посторонних частицах”, которых в препарате заведомо не было, что обеспечивалось специальным обеспыливанием. Этот никак не прокомментированный группой Р. Пекоры эффект сильно затруднил ей попытки объяснить поведение ДНК с позиций казалось бы хорошо разработанной теории Цимма и Роуза для динамики полимеров в водных растворах. И это еще раз было подтверждено в Японии Матсумото и др. прямым наблюдением “аномально” броунирующей флуоресцентно-меченой ДНК. Представляется, что в работе группы Пекоры cветорассеяние происходило не только на реальных фрагментах ДНК, но и на волновых следовых структурах ДНК, оставляемых броунирующими молекулами этого суперинформационного биополимера в духе теории физического вакуума, где постулируется идея генерации фантомных торсионных аксионно-кластерных эквивалентов физических тел.
Что касается “аномалий” ДНК, обнаруженных в работе японцев, то здесь может иметь место также и вклад внешних физических полей, корригирующих квазиспонтанную динамику ДНК, вклад, который никак не принимался в расчет цитируемыми авторами.
ЗАПИСЬ ИК-ЛАЗЕРНОГО СИГНАЛА НА УРОВНЕ НЕЛИНЕЙНОЙ ДИНАМИКИ ДНК
Общая посылка данной части работы заключается в том, что хромосомный аппарат и его главная часть ДНК генерируют знаковые волновые структуры. Вместе с тем, геном способен на основе такого рода волновой памяти распознавать и корректировать пространственно-временную структуру биосистемы. Необходим простой и однозначный экспериментальный результат, который показал бы, что молекулы ДНК в принципе способны к памяти на внешнее электромагнитное поле. В качестве такового был выбран ИК-лазерный сигнал с учетом того, что ДНК in vivo оперирует таким излучением. Мы поставили несколько серий экспериментов для того, чтобы ввести in vitro такой искусственный лазерный сигнал в гель молекул ДНК с последующим анализом их нелинейной динамики как системы отображения ИК-лазерного воздействия на уровне явления возврата Ферми-Паста-Улама (ФПУ) [25]. Для введения такого рода сигнала в нелинейно-динамический континуум геля ДНК мы использовали импульсный режим работы ИК-лазера Ga-As с длиной волны 890 нм, частотой повторения импульсов 600 Гц со средней мощностью (минимум 0,8; максимум 3,1) Вт с временем однократной экспозиции 4 сек. Регистрацию воздействий лазера и подготовку образцов ДНК из эритроцитов кур вели в соответствии с [25], в частности, с использованием метода корреляционной лазерной спектроскопии. Анализ поведения временных автокорреляционных функций (АКФ) светорассеяния ДНК показал, что сигнал ИК-лазера запоминается биополимером в форме периодической стохастизации АКФ и носит долговременный и устойчивый характер. Периодические повторы стохастических АКФ допустимо трактовать как одну из форм явления возврата Ферми-Паста-Улама, сочетанного со свойственной этому явлению памятью. Замораживание ДНК геля в течение недели не влияет на приобретенную память на ИК-лазерный сигнал. После размораживания периодическая стохастизация АКФ данного препарата сохраняется, если поддерживать препарат в высокополимерной форме. Таким образом, удалось впервые осуществить запись внешнего искусственного импульсного ИК-лазерного воздействия на уровне нелинейной динамики ДНК, что может служить простейшей реалистической моделью эпигеноволновых процессов in vivo.
О ВОЗМОЖНОСТИ СОЗДАНИЯ ЛАЗЕРА НА ИНФОРМАЦИОННЫХ БИОМАКРОМОЛЕКУЛАХ [30]
Прошло несколько десятилетий после того, как лауреаты Нобелевской премии академики РАН А.Н.Прохоров, Н.Г.Басов (Россия) и Чарльз Таунс (США), высказали идею, а затем реализовали ее, о возможности создания квантовых генераторов. Сейчас трудно сказать, в какой области науки и техники они не применяются (от биологии и медицины до лазерного термоядерного синтеза). Последующие исследователи внесли свой крупный вклад в развитие этой проблемы.
В данной части работы ставится вопрос: можно ли in vitro создать лазер на информационных биомакромолекулах, прежде всего на ДНК, РНК и хромосомах? Вряд ли может идти речь о создании энергетически мощных лазеров на этих структурах. Вопрос звучит по-иному: какие новые знания мы можем получить о ДНК, РНК и хромосомах, создав такой лазер и исследуя характер его излучения? Можно думать, что это будут принципиально новые данные. Например, об их нелинейной динамике, в том числе солитонного типа, о ровибронных колебаниях, о модуляциях дисперсии оптического вращения и кругового дихроизма, переносе энергии в другие, ранее недоступные (в таком варианте методологии) слои информации. При этом динамические модификации лазерного пучка могут иметь cемантико-гено-биознаковый характер и поэтому будут обладать мощной биологической активностью.
Первые соображения по этому поводу были предложены нами ранее [25,30]. В том числе обсуждалась идея о создании лазерной системы на Фрёлиховских модах [3]. Сложность доказательства правильности всех этих мыслей состоит в том, что большинство генетических структур, содержащих в своем составе ароматические и гетероциклические кольца, “прозрачны” для характерного спектрального диапазона l @ 350-400нм. Трудность также и в том, что если использовать мощную оптическую накачку, то это, учитывая “хрупкость” биоструктур, неизбежно приведёт к их разрушению.
В настоящей главе для реализации некоторых из обсуждавшихся положений проведено исследование in vitro спектров двухфотонно-возбуждаемой люминесценции (ДВЛ) геле-жидкокристаллических препаратов нуклеогистона, являющегося суммарной фракцией хромосом, в которой преобладают гистоновые белки, и ДНК (стандартные высокополимерные препараты фирмы “Sigma”). Для существенного увеличечения интенсивности ДВЛ генетических структур нами предложен способ активации люминесценции за счет введения в состав исследуемых образцов активаторов (доноров) ДВЛ-определенных (близких по спектру оптического поглощения ДНК и нуклеогистону) органических молекул. Такие молекулы характеризуются большой интенсивностью спектров излучения, которые располагаются в области собственного оптического поглощения ДНК и нуклеогистона. В качестве активатора мы использовали кристаллический препарат димедрола, структура которого включает пару бензольных колец. Для димедрола это обеспечивает интенсивный спектр ДВЛ, имеющий вид широкой асимметричной полосы в диапазне 280 - 350нм.
Для фотонной накачки исследуемых препаратов мы применяли лазер на парах меди. Этот лазер работает в стандартном импульсно-периодическом режиме с частотой следования импульсов 10кГц, со средней мощностью
![]() Вт, пиковой мощностью 10
Вт, пиковой мощностью 10![]() Вт, длинами волн генерации l = 510,8нм и 578,2нм (зеленая и желтая линии), длительностью импульсов
Вт, длинами волн генерации l = 510,8нм и 578,2нм (зеленая и желтая линии), длительностью импульсов
![]() нс. Лазерное излучение направляли на исследуемый образец в виде сфокусированного пятна размером
нс. Лазерное излучение направляли на исследуемый образец в виде сфокусированного пятна размером ![]() мм. Применение такого лазера как инициатора ДВЛ оказалось весьма эффективным при изучении электронно-колебательных спектров белков, ДНК, нуклеогистона и их компонентов (пурины, пиримидины, аминокислоты [19,30]). Регистрирующая аппаратура включала: фильтр для выделения лазерных линий с l =510,8 и 578,2 нм, фильтр для выделения излучений люминесценции в УФ и фиолетовом диапазонах (с подавлением лазерного излучения), монохроматор (тип МДР-2) для сканирования спектра в широком интервале (от УФ до видимой области), двухкоординатный самописец для регистрации спектров, измеритель для контроля опорного сигнала и определения эффективности наблюдаемого сигнала. Для подавления тепловых шумов применяли строб-импульс длительностью 25-30нс, синхронизированный с импульсом возбуждения. Регистрацию вторичного импульса излучения проводили с ФЭУ-130. Исследования спектров ДВЛ геле-жидкокристаллического препарата ДНК в смеси с димедролом (ДНК-ДЛ) и нуклеогистона с димедролом (НГ-ДЛ) показали, что амплитуда ДВЛ спектра ДНК-ДЛ лишь на порядок меньше таковой спектра ДВЛ чистого димедрола. Это обеспечивает существенное увеличение интенсивности ДВЛ смеси ДНК-ДЛ по сравнению с чистым препаратом ДНК в форме жесткого геля [19]. На этом же спектре обнаруживается ряд дополнительных особенностей изучаемых смесей. Оказалось, что квантовый выход ДВЛ для смеси НГ-ДЛ ниже, чем для смеси ДНК-ДЛ. Другая характерная черта - разгорание или тушение ДВЛ во времени. Для НГ-ДЛ наблюдается нарастание ДВЛ во времени. Обратный эффект наблюдается в случае ДНК-ДЛ. Представляет интерес присутствие вибронной структуры в спектрах ДВЛ в виде отдельных перекрывающихся полос в области 310-370 нм, особенно для ДНК-ДЛ. Такая структура близка к ранее наблюдавшимся спектрам ДВЛ для нуклеозид-трифосфатов [19].
мм. Применение такого лазера как инициатора ДВЛ оказалось весьма эффективным при изучении электронно-колебательных спектров белков, ДНК, нуклеогистона и их компонентов (пурины, пиримидины, аминокислоты [19,30]). Регистрирующая аппаратура включала: фильтр для выделения лазерных линий с l =510,8 и 578,2 нм, фильтр для выделения излучений люминесценции в УФ и фиолетовом диапазонах (с подавлением лазерного излучения), монохроматор (тип МДР-2) для сканирования спектра в широком интервале (от УФ до видимой области), двухкоординатный самописец для регистрации спектров, измеритель для контроля опорного сигнала и определения эффективности наблюдаемого сигнала. Для подавления тепловых шумов применяли строб-импульс длительностью 25-30нс, синхронизированный с импульсом возбуждения. Регистрацию вторичного импульса излучения проводили с ФЭУ-130. Исследования спектров ДВЛ геле-жидкокристаллического препарата ДНК в смеси с димедролом (ДНК-ДЛ) и нуклеогистона с димедролом (НГ-ДЛ) показали, что амплитуда ДВЛ спектра ДНК-ДЛ лишь на порядок меньше таковой спектра ДВЛ чистого димедрола. Это обеспечивает существенное увеличение интенсивности ДВЛ смеси ДНК-ДЛ по сравнению с чистым препаратом ДНК в форме жесткого геля [19]. На этом же спектре обнаруживается ряд дополнительных особенностей изучаемых смесей. Оказалось, что квантовый выход ДВЛ для смеси НГ-ДЛ ниже, чем для смеси ДНК-ДЛ. Другая характерная черта - разгорание или тушение ДВЛ во времени. Для НГ-ДЛ наблюдается нарастание ДВЛ во времени. Обратный эффект наблюдается в случае ДНК-ДЛ. Представляет интерес присутствие вибронной структуры в спектрах ДВЛ в виде отдельных перекрывающихся полос в области 310-370 нм, особенно для ДНК-ДЛ. Такая структура близка к ранее наблюдавшимся спектрам ДВЛ для нуклеозид-трифосфатов [19].
Механизм резкого увеличения квантового выхода ДВЛ нуклеогистона и ДНК при наличии донор-активатора (димедрола) может быть объяснен быстрой квазирезонансной передачей энергии от возбужденных молекул димедрола к исследуемым геноструктурам. Наблюдаемая при этом тонкая многополосчатая структура ДВЛ спектров коррелирует с характером вибронных полос для ряда ароматических и гетероциклических соединений, включая чистые нуклеозид-трифосфаты и ДНК [19]. Возникновение такого рода дискретизации спектров можно трактовать переходом электронов биомакромолекул с электронного терма S1 на возбужденные колебательные уровни основного состояния S0. В связи с этим может быть реализована инверсная заселенность на переходах ![]() при достаточном заселении терма
при достаточном заселении терма ![]() .
.
Проведем оценки необходимой интенсивности ![]() лазерного излучения для создания инверсии (суперфлуоресценции) в условиях проведенных опытов.
лазерного излучения для создания инверсии (суперфлуоресценции) в условиях проведенных опытов.
Условия инверсии записываются следующим образом:
![]() , (1)
, (1)
где ![]() - плотность рабочих молекул в состоянии
- плотность рабочих молекул в состоянии ![]() ,
, ![]() - плотность молекул в состоянии
- плотность молекул в состоянии ![]() ,
, ![]() и
и ![]() - соответствующие статистические веса квантовых уровней.
- соответствующие статистические веса квантовых уровней.
Плотность заселенности оценивается из соотношения
![]() , (2)
, (2)
где ![]()
![]() - скорость заселения уровня
- скорость заселения уровня
![]() ,
, ![]() - скорость его распада за счет излучательного процесса и (или) безызлучательных процессов.
- скорость его распада за счет излучательного процесса и (или) безызлучательных процессов.
Для величины
![]() имеем оценку:
имеем оценку:
![]() (3)
(3)
где W и
![]() - энергия и длительность лазерного импульса,
- энергия и длительность лазерного импульса,
![]() - эффективный объем среды, в котором реализуется двухфотонное поглощение (S - площадь поперечного сечения сфокусированного светового пучка, падающего на исследуемый образец,
- эффективный объем среды, в котором реализуется двухфотонное поглощение (S - площадь поперечного сечения сфокусированного светового пучка, падающего на исследуемый образец,![]() - эффективная длина проникновения излучения в образец),
- эффективная длина проникновения излучения в образец),![]() - плотность биомакромолекул (ДНК или нуклеогистона).
- плотность биомакромолекул (ДНК или нуклеогистона).
С учетом соотношений (1) - (3) условие для cоздания инверсной заселённости суперфлуоресценции записывается в виде
![]() .
.
Используя характерные данные
![]()
![]() (для
(для![]() нм) =
нм) =
![]() Дж,
Дж,
t @ 10нс,![]() ,
,![]()
![]() ,
,
получаем оценку
![]() ,
,
что близко к использованным значениям интенсивности в наших экспериментах.
Проведенные экспериментальные исследования и их теоретические оценки дают основание достаточно уверенно предполагать, что при используемых режимах двухфотонного возбуждения с использованием активатора-димедрола в геноструктурах in vitro реализуется усиление люминесценции, т.е. излучение ДНК и нуклеогистона носит характер суперфлуоресценции.
Не исключено, что в биосистеме роль димедролоподобных веществ в качестве активаторов могут выполнять эндогенные соединения, прямо или косвенно взаимодействующие с ДНК и хромосомами (стероидные гормоны, углеводы, нуклеозид -моно, -ди и -трифосфаты, некоторые витамины (например, рибофлавин), ароматические и гетероциклические аминокислоты, катехол- и индолалкиламины, некоторые антибиотики, наркотические вещества (например, эндогенные морфины - метаболиты этанола и пептиды-эндорфины), алкалоиды, токсины, ко-факторы ферментов, гем-содержащие белки и другие многочисленные органические соединения, содержащие бензольные и гетероциклические компоненты.
Неясны условия реализации инверсной электронной заселенности геноструктур in vivo, близкие тем, которые использовались нами в режимах ДВЛ. Такие условия могут создаваться в биосистемах, например, за счет фотон-фононных взаимодействий в ДНК в рамках теории Дике.
Однако, это относится к чисто физическим механизмам. Что касается физико-биохимических процессов, приводящих к лазерной накачке ДНК и хромосом in vivo, то в качестве таковых можно предсказать наличие в биосистемах мощных АТФ-азных систем, поставляющих энергию для перевода генетических структур в биокогерентные состояния (аналогичные тем, что как частный случай изложены в настоящей главе).
ТЕОРЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ ВОЗМОЖНОСТИ СОЗДАНИЯ БИОЛАЗЕРА НА ФРЕЛИХОВКИХ МОДАХ [3]
В данной главе обсуждается и аналитически рассматривается возможность создания перевозбужденного состояния основной (выделен-ной) коллективной Фрелиховской моды за счет когерентного резо-нансного взаимодействия электромагнитного (амплитудно-модулиро-ванного) излучения с Фрелиховским осциллятором. В рамках по-нятий лазерной физики речь идет о создании инверсной заселенности между квантовыми уровнями выделенной колебательной моды и, в итоге, о реализации “in vitro-in vivo” суперфлуоресценции и лазерной генерации с использованием в качестве рабочих тел молекул ДНК, РНК, белков, а также таких надмолекулярных структур, как рибосомы, полирибосомы и хромосомы.
Подчеркнем, что в отличие от Фрелиховского подхода, в котором подразумевается квазинеравновесное состояние (колебательная температура выделенной моды превосходит таковую “тепловой бани” Tvib>Teq>0, т.е. колебания квазиравновесны), в данной работе оценены условия, при которых система рассматриваемых биосубстратов инвертирована (Tvib=1/2![]() ,
, ![]() >=1/2
>=1/2![]() , (18)
, (18)
так что
![]()
Эффект инверсии (![]() реализуется при условии
реализуется при условии
![]() . (19)
. (19)
Если параметр глубины модуляции ![]() лежит в диапазонах, где значения функции Бесселя
лежит в диапазонах, где значения функции Бесселя ![]() отрицательны, то реализуется режим перевозбуждения системы (информационных биомакромолекул и надмолекулярных структур).
отрицательны, то реализуется режим перевозбуждения системы (информационных биомакромолекул и надмолекулярных структур).
Таким образом, высказана идея принципиальной возможности создания биолазеров на Фрелиховских модах in vitro, а также инициации таких процессов в живой клетке в дополнение (или коррекции) к известным естественным лазероподобным процессам в биосистемах. Показано, что в определенных условиях - в случае когерентного (резо-нансного) взаимодействия амплитудно-модулированного внешнего электромагнитного излучения с Фрелиховской модой - система информационных биоструктур может существовать в перевозбужденном состоянии, что является необходимой предпосылкой для создания знаконесущих биолазеров.
Необходимо отметить,что описанный выше механизм формирования биолазеров на основе молекул ДНК позволяет подойти к попытке реализации еще одной фундаментальной гипотезы Фрелиха о возможности перекачки энергии kТ внутриклеточной жидкости в энергию электрических колебаний в молекуле ДНК. В соответствии с этой гипотезой стохастические тепловые колебания kТ раствора могут резонансно взаимодействовать (в определенном интервале частот) с колебательными модами молекулы ДНК, и благодаря тому, что как молекула ДНК, так и молекулы белков представляют собой распределенные нелинейные колебательные структуры, часть энергии может группироваться в низкочастотных модах этих молекул. Иными словами, молекула ДНК в растворе может частично преобразовывать энергию колебаний kТ в энергию собственных мод. Заметим, что даже в рамках предложенного квазили-нейного подхода проблема перекачки тепловой энергии раствора может быть сведена к механизму затухания квантового осциллятора, который был предложен А.Пиппардом. C учетом этого в уравнение Шредингера вводится комплексный потенциал, интерпретирующий передачу энергии осциллятора большому числу мод расширяющегося сферического резонатора. Если размеры этого резонатора конечны, как в случае с живой клеткой, то возникнет резонансный обмен энергии между модами kТ раствора и электрическими модами молекулы ДНК. Эти рассуждения также говорят в пользу того, что и в водно-жидкокристаллическом электролите клеточно-тканевого пространства биосистемы генетические молекулы могут функционировать как биолазеры.
Надо указать на существенное обстоятельство относительно принципиальной возможности реализации возбуждения Фрелиховских мод “in vitro” по биохимическому пути, а именно за счет энергии гидролиза АТФ и других нуклеозид-трифосфатов, а также за счет других макроэргических соединений живой клетки. В данном случае мы будем искусственно повторять то, что эволюционно и (или) иным путем дано биосистемам как основная информационная и, может быть, энергетическая фигура. Эта часть наших исследований ставит определенные нравственные и этические проблемы применения биолазеров.
АНТЕННАЯ МОДЕЛЬ ФИЗИКО-МАТЕМАТИЧЕСКИЙ ФОРМАЛИЗМ [16]
Как уже неоднократно отмечалось, функционирование ряда биологических макромолекул (в частности, ферментов) и других биологических соединений во многом определяется процессами, происходящими в активных центрах, окруженных биополимерными цепочками, имеющими знаковую топологию. Исходя из такого представления о структуре информационных биомакромолекул, естественно предположить, что их взаимодействие с физическими полями внешних по отношению к биосистеме и внутренних (организменных) излучений приводит к возбуждению дипольно-активных колебаний мономеров, формирующих указанную цепочку, а те, в свою очередь, индуцируют колебания в активном центре. Иными словами, такая система будет работать как своеобразная антенна. Эти возбужденные колебания способны привести к переходу биомакромолекулы в другое конформационное (топологическое, знаковое) состояние.
Подобная концепция в принципиальном плане адекватна целому ряду функционально высокозначимых биомакромолекул, например, хлорофилла, гемоглобина, миоглобина и т. д. Эти макромолекулы объединяются двумя структурными качествами: 1) в их геометрическом центре расположен ион (в случае хлорофилла - ион магния, в случае гемоглобина - ион железа); 2) около иона симметрично расположены 4 пиррольных кольца (псевдоплоская структура).
Другими типами биополимеров, соответствующих антенной модели, могут быть cравнительно простые циклы типа валиномицина (переносчик ионов калия) и сложные надмолекулярные структуры хромосом, ДНК которых содержит высокоорганизованные ассоциаты таких металлов, как магний, кальций, никель, кобальт, медь, железо, цинк и др. При этом роль их неясна и сводится исследователями, в основном, к нейтрализации ОН-групп остатков фосфорной кислоты полинуклеотида. Представляется, что функции металлов в ДНК и РНК существенно более широкие и реализуются по линии знакового и (или) энергетического взаимодействия с эндогенными и экзогенными по отношению к биосистеме физическими полями. То же относится и к белкам, не содержащим порфириновый центр, но специфическим образом связывающим металлы. Например, таковыми можно считать сайт-специфические белки с доменами типа “цинковых пальцев”, участвующими в регуляции генов, подчас очень далеко отстоящих от этих управляющих белков. Атомы металлов ДНК и белков могут резонансно взаимодействовать по электромагнитным каналам в рамках понятий антенной модели. Еще раз обозначим понятие антенной модели.
Внешняя энергия (в частности, связанная с резонансным взаимодействием крайне высокочастотных электромагнитных излучений с белками) поступает на периферию, т. е. на ансамбль субъединиц (не обязательно идентичных по структуре). В результате активной “беседы”, предопределенной биохимическими связями, между периферийными акцепторами (получившими закодированную энергию) и центром-ассоциатом (в данном случае ионом металла гемсодержащих белков), последний получает энергию (информацию), что и вызывает биологическое действие. Степень реакционной способности биомакромолекул существенно зависит от уровня возбуждения центральных субъединиц. Рассмотрим в деталях потенциальные механизмы волновых взаимодействий физических полей и активных центров информационных биомакромолекул в рамках предлагаемой нами антенной модели.
В качестве простейшей модели для иллюстрации антенного эффекта рассмотрим двумерную замкнутую (циклическую) цепочку мономеров. В центре цикла расположен активный центр, связанный с мономерами цепочки диполь-дипольным взаимодействием.
Обозначим координатные смещения мономеров через
![]() , а смещение активного центра через
, а смещение активного центра через ![]() . Для потенциальной функции имеем:
. Для потенциальной функции имеем:
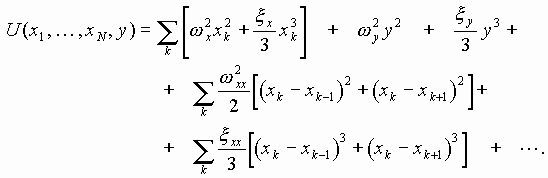 (1)
(1)
Первые два члена в (1) соответствуют колебаниям мономеров (второй член учитывает ангармонизм); последние два члена отвечают за связи между мономерами, Остальные члены отвечают за связи между мономерами и активным центром.
Уравнения движения запишем в виде:
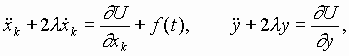 (2)
(2)
где ![]() - внешняя монохроматическая сила, действующая только на мономеры,
- внешняя монохроматическая сила, действующая только на мономеры, ![]() - коэффициент затухания, введенный феноменологически (простоты ради принят одинаковым и для мономеров, и для активного центра).
- коэффициент затухания, введенный феноменологически (простоты ради принят одинаковым и для мономеров, и для активного центра).
С учетом (1), система уравнений (2) приобретает вид:
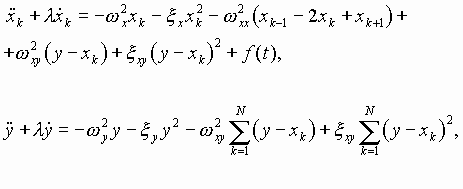 (3)
(3)
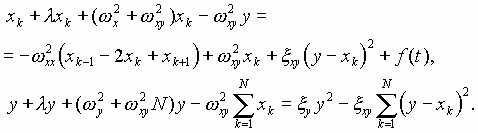 (4)
(4)
Введем общую координату для ансамбля мономеров
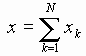 . (5)
. (5)
тогда система уравнений (4) в линейном приближении приобретает вид:
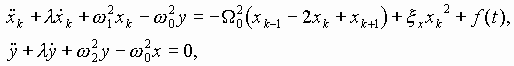 (6)
(6)
где:
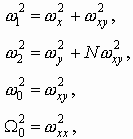
![]() - число мономеров.
- число мономеров.
С учетом (5) имеем ![]() (7.1)
(7.1)
![]() (7.2)
(7.2)
Из (7.2) следует
![]() (8)
(8)
Подстановка (8) в (7.1) дает
![]() .
.
(9)
Соответствующее характеристическое уравнение имеет вид (после подстановки ![]() в однородное уравнение):
в однородное уравнение):
![]() (10)
(10)
Обозначив ![]() имеем
имеем
![]()
так что
![]() (11)
(11)
В дальнейшем предполагается выполнение неравенств:
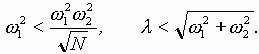 (12)
(12)
Первое условие соответствует случаю слабой связи между мономерами и активным центром, второе - малому затуханию мономерных осцилляторов.
Для собственных значений имеем
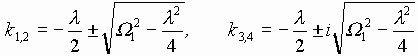 , (13)
, (13)
где введены коллективные частоты:
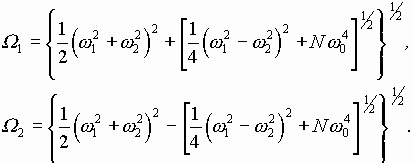 (14)
(14)
Нас интересуют вынужденные колебания (внешняя сила ![]() ):
):
![]() . (15)
. (15)
Подстановка (15) в (9) и приравнивание соответствующих коэффициентов при
![]() и
и ![]() дают систему алгебраических уравнений:
дают систему алгебраических уравнений:
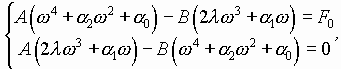
где:
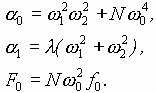
В результате получаем
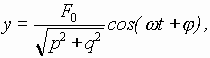
где 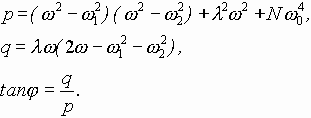
После несложных, но громоздких преобразований для вынужденных колебаний активного центра получаем:
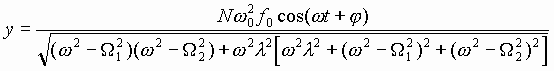 . (16)
. (16)
Из (16) видно, что наибольшая амплитуда вынужденных колебаний активного центра достигается в условиях коллективного резонанса: либо ![]() , либо
, либо
![]() .
.
В любом из этих случаев для амплитуды вынужденных колебаний имеем:
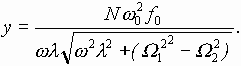 (17)
(17)
Из (17) следует, что наибольший эффект резонансной раскачки активного центра достигается при большем числе периферийных субъединиц “антенны”, при более высоком значении коэффициента связи активного центра с мономерами, при наименьшем коэффициенте затухания и при наименьшем дисбалансе коллективных мод.
Нетрудно определить и “хореографию” (динамику вынужденных колебаний) отдельных мономерных единиц. В соответствии с (6) уравнение для k -го мономера запишем в виде:
![]() (18)
(18)
Вводя коллективные координаты
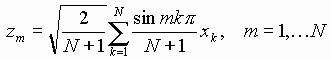
и применяя метод линейной алгебры, получаем для вынужденных колебаний мономеров:
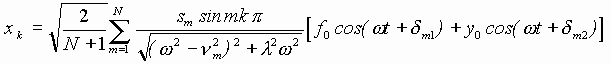 ,
,
(19)
где:
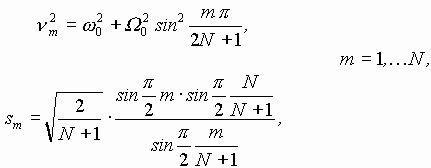
![]() - определяется из (16)
- определяется из (16)
Таким образом, в рамках антенной модели наибольший эффект воздействия внешнего монохроматического поля
![]() ре-ализуется в условиях коллективного резонанса:
ре-ализуется в условиях коллективного резонанса:
![]() .
.
Повторяя рассуждения раздела 2, можно сделать также следующие выводы:
1) При реализации амплитудной модуляции внешнего сигнала имеют место дополнительные возможности резонансного воздействия на биомакромолекулы на частотах:
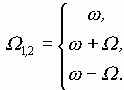
2) Учет нелинейности при квадратичной связи для монохроматического сигнала привносит дополнительный резонанс на второй гармонике ![]()
3) Учет нелинейности при амплитудной модуляции определяет еще ряд резонансных возможностей:
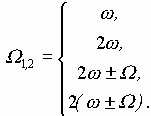
Таким образом, при действии резонансного электромагнитного поля на биомакромолекулы с активным центром, содержащим атомы металлов, существенную роль играют коллективные волновые эффекты. В этом случае свойства самого излучения предопределяют широкие возможности регуляторного влияния на динамику биомакромолекул в целом и, следовательно, на биопроцессы, в которых они принимают участие, тем самым прямо или косвенно реализуя управляющие и (или) дезорганизующие сигналы.
КОНВЕРСИЯ ЭПИГЕНОСИГНАЛОВ В ЭЛЕКТРОМАГНИТНЫХ СОЛИТОННЫХ СТРУКТУРАХ, ИХ ТРАНСПОЗИЦИЯ В ГЕНОМ БИОСИСТЕМ-АКЦЕПТОРОВ
Детально методы и эксперименты по дистантной трансляции и биологической активности электромагнитных солитонов, синтезированных на основе явления возврата Ферми-Паста-Улама (ФПУ) и промодулированных эпигеносигналами, приведены в работе автора [25]. Здесь же отметим принципиальные позиции, разграничивающие прежние представления о работе генов как чисто вещественных образований и наших представлений о знаковых волновых излучениях (“волновых генах”) хромосомного континуума.
Реальные и достоверные эксперименты в области волновой генетики первым начал проводить Дзян Каньджэн. Итоговые работы его известны [Дзян Каньджэн. 1993. Биоэлектромагнитное поле - материальный носитель биогенетической информации. Аура-Z. № 3. с.42-54. Патент №1828665 “Способ изменения наследственных признаков биологического объекта и устройство для направленной передачи биологической информации”. заявка № 3434801. приоритет изобретения 30.12.1981г., зарегистрировано 13.10.1992г.]. Прибор Дзян Каньджэна, дистантно (десятки сантиметров) передающий “волновые гены” от донора к реципиенту, использует собственные излучения биосистем-доноров, причем, как считает автор, только в СВЧ-дипазоне электромагнитных полей. Авторское теоретическое обоснование эффектов, полученных с помощью этой аппаратуры, откровенно слабо, а точнее, просто неверно. Однако результаты убедительны. Это “волновые” гибриды пшеницы и кукурузы, земляного ореха и подсолнуха, огурца и дыни, утки и курицы, козы и кролика. Полученные гибридами признаки передаются по наследству. Блестящий эмпирик Дзян Каньджэн оказался неспособным понять тонкие механизмы открытых им эффектов, но это нисколько не умаляет значимость результатов, суть которых в доказательстве реальности “волновых генов”.
Вслед за этими исследованиями мы, уже своими методами, подтвердили принципиальную возможность дистантной трансляции и акцепции эпигенетических управляющих сигналов in vitro-in vivo в форме особого вида электромагнитного поля. Это еще раз подтвердило идеи А.Г. Гурвича, А.А. Любищева и В.Н. Беклемишева, но на современном уровне. Стало ясно, что “волновые гены” могут существовать, в частности, как одна из форм явления возврата ФПУ, что хорошо коррелирует с нашими данными по ФПУ-возврату на уровне нелинейной динамики ДНК in vitro. Именно это фундаментальное явление и легло в основу конструкции генератора ФПУ, приближенно моделирующего знаковые электродинамику и акустику ДНК in vivo и потому способного “считывать” и ретранслировать управляющие метаболизмом биосистем солитонные структуры с хромосомного континуума биосистем-доноров и резонансно вводить их в геном биосистем-акцепторов.
В связи с принципильной важностью феномена моделирования ФПУ-процессов в геноме высших биосистем при помощи особых радиоэлектронных устройств (ФПУ-генераторов) имеет смысл остановиться несколько подробнее на феномене ФПУ-возврата. Это явление было обнаружено в 1949 г. как результат компьютерного исследования динамики колебаний в цепочках нелинейно связанных осцилляторов. Оказалось, что против всякого ожидания энергия первоначального возмущения крайних осцилляторов в таких цепочках не термолизовалась, а распределившись по высшим гармоникам, затем вновь собиралась в спектр первоначального возмущения. При увеличении числа осцилляторов в цепочке картина возврата энергии неизменно сохранялась. Эта проблема получила название возврат Ферма-Паста-Улама по именам Э.Ферми, Д.Паста и З.Улама, которые первыми исследовали эту задачу. В дальнейшем возврат ФПУ был экспериментально обнаружен в длинных электрических линиях с нелинейными элементами в плазме, а также в динамике волн на глубокой воде. Замечательным свойством возврата ФПУ оказалось наличие “памяти” в его спектре к начальным условиям его активных мод.
Результаты исследований в области изучения возврата ФПУ позволили теоретически рассмотреть молекулу ДНК в виде электрического резонатора ФПУ. В этой модели динамика волны плотности электронов, распространяющейся вдоль сахаро-фосфатных цепей молекулы ДНК, рассматривалась в рамках нелинейного уравнения Шредингера в форме, предложенной Юэном и Лэйком для описания динамики солитонных волн на глубокой воде. При этом осцилляции плотности электронов в структурах нуклеотидов понимали как возмущающие точечные источники, расположенные на одинаковых расстояниях вдоль сахаро-фосфатных цепочек ДНК, интерпретируемых как длинная электрическая линия.
В дальнейшем эта модель была развита А. А. Березиным совместно с автором [25]. В частности, были рассмотрены электрические поля (E', E") обеих цепочек ДНК, где E' - средняя амплитуда напряженности электрического поля за один пространственный период стоячих волн в первой цепи ДНК, а E" - средняя амплитуда напряженности электрического поля за один временной период стоячих волн во второй цепи. Если принять, что колебания E' и E" генерируются молекулой ДНК в окружающее пространство, тогда вне молекулы ДНК поля E' и E" образуют сферические фронты. При этом в силу представления стоячих волн в молекуле ДНК в виде двух противоположно направленных бегущих фронтов возмущений, от источника (молекулы ДНК) будет расходиться сферическая волна E', а к источнику будет сходиться сферическая волна E", поскольку волны от молекулы излучаются в нелинейную среду - внутриклеточную жидкость. Динамика этих волн может быть описана в сферических координатах. Для E" частное решение будет выглядеть аналогично. Было получено выражение, представляющее собой интенсивность электрической волны на сфере определенной толщины вокруг молекулы ДНК, своего рода “сферическая голограмма”, существующая в электролите клеточно-тканевого пространства в сферическом слое. Предложенная модель указывает на возможность существования вокруг молекулы ДНК в составе хромосом сферических акустико-электромагнитных солитонов (бри-зеров), которые интегрально отображают структуру хромосомного континуума и могут двигаться за пределы клеточных ядер или совершать колебательные движения относительно некоего положения равновесия и которые содержат статико-динамические квазиголографические (в общем случае дифракционные) решетки с эпигенознаковой образно-семан-тической нагрузкой. Такие решетки отображают текущее и (или) относительно постоянное пространственно-временное состояние организма в каждой области многомерной структуры высших биосистем, где в данный момент находится бризер. Наличие тепловых возмущений (kT) молекулы ДНК, а также возможность существования фуранозных колец нуклеотидов в виде двух конформаций, приводят к усложнению модели и необходимости введения в нее фазовых флуктуаций электронной плотности.
Однако, учитывая, что спектр ФПУ может служить преобразователем стохастических колебаний в детерминированные, стохастическая компонента динамики колебаний электронной плотности в молекуле ДНК является, вероятно, ее атрибутом.
ГЕНЕРАТОР ПАКЕТОВ УЕДИНЕННЫХ ВОЛН (СОЛИТОНОВ) В ФОРМЕ ВОЗВРАТА ФЕРМИ-ПАСТА-УЛАМА
Данная теоретическая модель нелинейной знаковой акусто-электродинамики ДНК легла в основу создания семейства радиоэлектронных устройств - генераторов пакетов уединеных волн в форме возврата Ферми-Паста-Улама (ФПУ-генераторов), предназначенных для генерации электромагнитных волн (солитонов), обладающих характерной пространственно-временной структурой возврата ФПУ, которое выражается в периодическом переходе колебательной структуры от упорядоченного состояния к хаотическому и обратно. При этом в упорядоченном состоянии первоначальная форма волнового пакета и его пространственно-временной спектр полностью повторяются. Важной особенностью ФПУ-генераторов является пространственно-временная структура его поля, которая является относительно простой физической моделью колебательной структуры молекулы ДНК. Это свойство позволяет использовать генератор в экспериментах по исследованию собственных колебаний в препаратах ДНК и по информационному взаимодействию биологических систем, о которых говорилось выше. Первые модели таких генераторов были созданы А. А. Березиным и соавторами (1988, 1989 г. г. ), а затем в 1991г. были принципиально дополнены П. П. Гаряевым и Г. Г. Комиссаровым за счет интеграции в их схемы кодирующего акустического ввода.
Принципиальная схема генератора содержит ФПУ-резонатор в виде двух длинных линий с подключенными к ним нелинейными элементами (туннельные диоды). Напряжение смещения туннельных диодов задается стабилизаторами на транзисторах и стабилитроне. Выбор рабочей точки туннельных диодов и способ их подключения к ФПУ-резонатору обеспечивают форму и спектр колебаний генератора, которые соответствуют нормальным колебаниям одномерной решетки слабо связанных нелинейных (ангармонических) осцилляторов с периодическими граничными условиями, при которых наблюдается явление возврата ФПУ. Для модуляции поля генератора внешними акустическими сигналами может быть использован угольный микрофон. Генератор питается от двух аккумуляторов типа ЦНК 0, 45-I-У2.
С помощью ФПУ-генератора и эмбрионов-доноров удалось непермиссивно дистантно (20 см - 2, 0 м) осуществить эмбриональную индукцию нейральных и мезодермальных производных в ткани эктодермы ранней гаструлы шпорцевой лягушки. Были получены результаты и по восстановлению нативной структуры у аберрантных радиационно поврежденных хромосом пшеницы и ячменя [25,29]. Это показывает реальность существования и моделирования знаковых электромагнитных полей геноволнового уровня, управляющих стратегическим метаболизмом биосистем, их наследственностью, и подтверждает близкие результаты, полученные Дзян Каньджэном.
ЕДИНСТВО ФРАКТАЛЬНОЙ СТРУКТУРЫ ДНК-"ТЕКСТОВ" И ТЕКСТОВ НА ЕСТЕСТВЕННЫХ ЯЗЫКАХ
Существует и другая семантическая ниша знаковых процессов в наследственном аппарате высших биосистем, связанная с его квази-речевыми характеристиками, а также с генетической атрибутикой словообразований в естественных человеческих языках. Ранее получены доказательства, что развитие языков и человеческой речи подчиняется законам формальной генетики. По сути, “тексты” ДНК (квазиречь) и письменность людей, их разговор (истинная речь) выполняют одинаковые управленческие, регуляторные функции, но в разных фрактально-сцепленных масштабированиях. ДНК генетически функционирует на клеточно-тканевом уровне, а человеческая речь, как макрогенетическая структура, используется на уровне общественного суперорганизма. Нам удалось несколько отойти от предшествующей метафоричности использования понятий лингвистики применительно к ДНК, когда произвольно используют термины “слово”, “текст”, “пунктуация”, “грамматика”, интуитивно пытаясь понять иные измерения генома. Такому отходу способствовало применение теории фракталей и метода перекодировок к последовательностям ДНК и структуре текстов людей. Выяснилось, что ДНК и человеческая речь (тексты) обладают стратегически близкой фрактальной структурой в геометрическом смысле. Вероятно, это каким-то образом коррелирует с фрактальной структурой солитонного акустического и электромагнитного ФПУ-поля, генерируемого хромосомным аппаратом высших биосистем. Возможно, именно по этой причине нам удалось зарегистрировать управленческие эффекты на геномах растений, вызываемые с помощью ФПУ-трансформированной человеческой речи, которая резонансно взаимодействует с хромосомной ДНК in vivo [25,29].
Этот результат, осмысленный нами с позиций семиотико-волновой составляющей генетического кода, имеет существенное методологическое значение и для анализа таких суперзнаковых объектов, как тексты ДНК, и для генома в целом. Открываются принципиально новые смысловые ареалы хромосомного аппарата. Однако биологии и гено-лингвистике предстоит пройти еще большой путь, прежде чем картина знаковых рядов ДНК станет относительно ясной и понимаемой. Вводимый нами способ мышления относительно функций генома позволяет сопоставлять различные естественные последовательности ДНК и РНК с оценкой меры их сходства и различия, а также степени относительной сложности их знаковой структуры. И кроме того, что более важно, появляется метод сопоставления смысловых конструкций человеческой речи и кодирующих последовательностей ДНК. Если мы правы в своих логических и экспериментальных построениях, то в общем плане видны новые измерения в понимании мышления и сознания через их отображения в знаковых (смысловых) рядах на разных уровнях организации живой материи - на уровне человеческой речи (высшая форма сознания) и квазиречи генетических молекул (квази-сознание генома). Это хорошо соответствует математико-лингвистической модели Хомского, постулирующей общие принципы, которые лежат в основе любого языка и которые объединяются в “универсальную грамматику”. Такая “универсальная грамматика”, по Хомскому, является врожденной, т. е имеет генетические детерминанты. Это чрезвычайно важное обстоятельство, которое еще раз фокусирует мысль на супергенетическом родстве знаковых структур ДНК и речевых образований человека. В какой-то мере мы подтвердили указанное положение, показав родство фракталей ДНК и человеческой речи. Хомский, вероятно, прав в том, что глубинные синтаксические конструкции, составляющие основу языка, передаются по наследству от поколения к поколению, обеспечивая каждому индивидууму возможность овладеть языком своих предков. То, что ребенок легко учится любому языку, объясняется как раз тем, что в своей основе грамматики всех языков совпадают. Суть человеческого языка инвариантна для всех людей. Можно думать, что эта инвариантность распространяется глубже, достигая макромолекулярных смысловых (“речевых”) структур хромосом. И этому есть определенные теоретико-экспериментальные подтверждения, полученные нами [25,29] и выводящие на существенно значимые методологические подходы мягкого регуляторного вхождения в неизвестные ранее семиотические пласты генетического аппарата высших биосистем. Но в этом же заключена и грозная опасность стратегических семиотико-волновых искажений знакополевого окружения Земли. Идеи волновой (и “речевой”) генетики находятся в фазе активного становления и поэтому необходима система жестко определенных запретов определенных экспериментов в этой области знания, подобная существующей в генной инженерии, например, по клонированию высших организмов.
Независимое подтверждение правильности гипотезы существования квази-вербального или, что одно и то же, образного уровня кодовых функций ДНК (в пределе хромосомного континуума биосистемы) может дать выход из ограниченного, а иногда неверного, функционального поля триплетного генетического кода, не объясняющего ни синтез белковых “текстов”, ни то, как в геноме зашифрована пространственно-временная структура организма. Конечная цель предлагаемого анализа выделение знаковых единиц различных уровней и понимание их семантики в функциональном пространстве ДНК- белок, которое, по крайней мере для ферментов, чрезвычайно гетерогенно (активный центр, сайты узнаваний, архитектоника водородно-гидрофобных сил самоорганизации пептидной цепи). Многоязычный метаболический “разговор” между информационными биополимерами клетки и их функционирование как результат обмена знаковыми биосигналами предполагают два взаимно коррелированных уровня этого обмена - вещественный и волновой. Вещественный хорошо изучен (матричное копирование ДНК-РНК-белки, взаимодействие антиген-антитело, самосборка клеточных структур), а тесно связанный с ним волновой уровень практически не изучен официальной наукой. И ситуация здесь непроста. Электромагнитные и акустические излучения белков, нуклеиновых кислот, мембран и цитоскелета хорошо известны. Представляется, что этот уровень информационных контактов клеточно-тканевого пространства выводит метаболические процессы в полевое измерение со своей “языковой” спецификой и регуляцией.
Рассматриваемые биоинформационные потоки, сцепленные с обменом веществ и энергии, не ограничиваются делением знаковых рядов на вещество и поле, но многократно умножаются фрактальностью этих рядов. Например, в акустико-электромагнитной компоненте сигнальных функций ДНК наблюдается фрактальность солитонного поля, формально описываемого уравнениями в рамках явления возврата Ферми-Паста-Улама. Это еще более усложняет семантический анализ белково-нуклеиновых и иных информационных контактов биоструктур. Можно полагать, что в живых клетках существует иерархия вещественно-волновых знаковых структур, где условную градацию “буква (фонема) - морфема - слово - предложение...” задает фрактальность этих структур. И то, что в одном масштабе является “предложением”, в другом, более крупном, может быть лишь “словом” и т. д. Другая сложность связана с понятием “рамки считывания”. Сдвиг на одну букву (или эквивалентное этому небольшое изменение фазы, поляризации, частоты физических полей в пространстве-времени биосистемы) может полностью поменять смысл читаемого текста (воспринимаемого образа), не говоря уже о том, что сами тексты, к примеру, в одних и тех же последовательностях ДНК могут быть записаны разными языками. Более того, нет запрета на понимание “текстов” жидкокристаллических хромосомных ДНК, как читаемых в трех- или n-мерном пространстве, когда “буквы слов” выстраиваются не только в одну линию и в одном измерении, но “читаются” вдоль и поперек, вверх и вниз и так далее. В таком процессе поочередно создается и уничтожается бесконечный континуум анизотропных “нитей текстов”, идущих во всех направлениях динамичного интерфазного хромосомного континуума всего пространства биосистемы. Предлагаемая логика неизбежна, если мы хотим понять сущность феномена жизни. Сказанное не следует рассматривать как предтечу пересмотра только принятой модели триплетного генетического кода. Она, эта модель, удобна,но только как исходная позиция, когда дешифрован (неточно и не до конца) первичный уровень кодонов иРНК, уровень вещественно-матричных геносигналов, составляющих 1 - 5% от всей массы геномной ДНК. Оставшаяся большая часть ДНК, существующая в понимании большинства генетиков в качестве "мусорной", несет, вероятно, стратегическую информацию о биосистеме в форме потенциальных и действительных волновых сигналов солитонной, голографической и иной образно-знаковой, в том числе и рече-подобной структуры (подробно см. выше главу “Пересмотр модели генетического кода”).
Вероятно, в прямой связи со всеми рассмотренными “аномальными” свойствами генома высших биосистем стоит феномен особого рода, требующий пристального внимания. Это проблема происхождения жизни, и в частности на Земле. Обсуждается она давно. Предположений много. Мы придерживаемся гипотезы панспермии, но не в том варианте, что на Землю были занесены некие споры-родоначальники всех жизненных форм. Вероятно, процесс естественной эволюции абиогенно возникшего “первичного бульона” из органических молекул - предшественников РНК, ДНК, белков и других существенных компонентов биосистем был сочетан с актом введения экзобиологической информации в первые нуклеиновые кислоты, она была артефактом. И эта информация была рече-подобной. “В начале было слово...”. И эти слова были фрактальны, условно начиная с дуплетно-триплетного кода ДНК-РНК, на первых этапах являющегося простейшим языком с четырех буквенной азбукой. Далее произошла трансляция в 20-буквенную азбуку белков и в более высокие языки в духе обсуждавшихся идей. Вообще гипотеза артефакта первичного языка ДНК широко обсуждается, начиная с пионерской работы В. И. Щербака, показавшего искусственность (привнесенность извне) коллективных симметрий генетического кода, вероятность эволюционного происхождения которых близка к нулю. Можно солидаризироваться с такой позицией не только по причине ее красоты и изящного способа доказательств, где в качестве реперных единиц теоретического анализа используются такие параметры, как нуклонные соотношения в аминокислотах и вырожденность генетического кода, но и потому, что она хорошо соответствует нашему мышлению. Однако, введем поправку. Поскольку на самом деле генетический код, то есть код биосинтеза белков, существенно отличается от принятого в начале 60-х г. хх века (см. выше), то и концепция артефакта кода также нуждается в уточнении. Можно предсказать в истинном (фрактальногетеромультиплетном) коде наличие и других знаковых математических образований, фрактально увеличенных по сравнению с теми, что открыл В.И. Щербак.
Развивая эту мысль и ранее выдвинутые нами идеи, скажем, что было бы наивным упрощением считать “языки” и “письменность” (“речь”) ДНК полным аналогом вербальных построений человека. Точнее будет полагать, что функции ДНК основаны прежде всего на ее метаязыке, являющимся грамматикой генома. Здесь чрезвычайно полезен анализ метаязыков А.Соломоником. Математика - тоже метаязык, он же и свод правил построения ее текстов. В отличие от обычной речи, в которой фразу с определенной мыслью можно сконструировать десятками разных способов, в математике ее вербальные (знаковые) ряды генерируются с помощью малого количества жестких правил. И они, правила, позволяют в автоматическом режиме получить предсказательный результат, как в нашем случае с антенной моделью, “предугадывающей” характер резонансных взаимодействий физических полей с информационными биомакромолекулами. На этом примере видно, как в конечном пункте математических метаязыковых (грамматически ориентированных) преобразований получается результат в форме физико-математического образа потенциального поведения важнейших компонентов биосистемы в ее полевом окружении и внутренней наполненности волновыми процессами. Хромосомы, возможно, также оперируют метаязыками для создания “идеальной” (физико-химико-математической) модели биосистемы как практически недостижимого прообраза реального организма. И такая модель будет более информативна по сравнению, например, с голографической моделью, и будет дополнять последнюю.
Если ДНК, хромосомы организмов Земли действительно являются одновременно донорами и акцепторами не только собственных волновых команд, но и неких внешних (возможно, экзобиологических) регуляторных волновых влияний, что было показано нами ранее, то новый искусственный, создаваемый людьми, электромагнитный семиотический канал вхождения в ноосферу и генофонд планеты Земля требует сверхвнимания в отношении уровня разумности и целесообразности наших, по сути неконтролируемых, супергенетических манипуляций. В этом случае мы будем входить в конкуренцию с вероятным экзобиологическим контролем. Полезно ли это и нужно ли? Сейчас ясного ответа нет. Возможно, мы вошли в бифуркационную вилку выбора стратегии эволюции человечества - или идти дальше по техногенному пути, или учиться мудрости у собственного тела, в котором сосредоточена мудрость Творца.
В качестве иллюстрации предложенного нами метода фрактального представления естественных и генетических текстов приведены матрицы плотности для текста на английском языке (руководство по программированию) и “текста” гена казеина (Cazein). Этот метод дает принципиально иную возможность количественного и качественного сравнения естественных и генотекстов. Аналогичный результат можно получить по- иному, и также новым методом, как это показано на графиках гистограмм сходства и различия для фланков и интронов большой группы генов. Таким же путем получена гистограмма сравнения естественных текстов для монографии автора “Волновой геном” и рассказов Ф.Абрамова [Неопубликованные результаты совместных исследований в соавторстве с М.Ю. Масловым (Математический институт РАН)].

Матрица плотности хаотически-игрового представления нуклеотидной последовательности (ген) в алфавите (A,T,G,C), кодирующей первичную структуру казеина (белок молока).
Рис. 11

Матрица плотности хаотически-игрового представления текста на английском языке (руководство по компьютерному программированию). Рассматривалась структура появления в тексте четырёх частей речи. Левый ближний угол соответствует слову “the”, правый ближний - слову “in”, левый дальний - “on”, правый дальний - ”of”.
Рис. 12

Сравнение фланков с интронами
Рис. 13
Сравнение монографий:Гаряев П.П. Волновой геном. М.,1994.
и Абрамов Ф. Были небыли. Рассказы. М., 1993.
Рис. 14
О ВОЗМОЖНОСТИ СОЗДАНИЯ БИОКОМПЬЮТЕРА НА ГЕНЕТИЧЕСКИХ СТРУКТУРАХ
В международном компьютерном еженедельнике “Сomputer World” (№ 5 от 3 октября 1995 г.) в рубрике “Подробности” была опубликована подборка статей, посвященная работам по созданию биокомпьютера на главной генетической молекуле - на ДНК. Томас Хоффман в статье “Болотная электроника...” описывает первые робкие попытки использования информационных биомакромолекул - некоторых белков (бактерио-родопсин, родопсин) в качестве субстратов записи-считывания информации как аналогов оптической дисковой памяти. Без сомнений, это интересное оригинальное направление, однако в данном случае ничего принципиально нового предложено не было, поскольку неважно откуда взято вещество-субстрат записи-считывания информации, на котором получают спектральные выжигания типа двоичного кода или с помощью лазеров записывают трехмерные изображения предметов в форме голограмм. Такое вещество может иметь абиогенное происхождение или, как в случае с родопсинами, извлекаться из биомембран солончаковых бактерий. В связи с этим, учитывая наши исследования, было бы логичным рассматривать молекулы ДНК как неразрывное единство Вещества и Поля также и в аспекте их участия как основной рабочей фигуры в искусственных биокомпьютерах. Это было бы полезно в развитии вычислительной техники и может привести к полной смене ее элементной базы в ряду: аналоговый-цифровой-“образный” или смысловой компьютер на ДНК.
Весной 1995 г. Леонард М. Адлеман, профессор вычислительных наук из Университета Южной Калифорнии, описал в журнале “Science” алгоритм использования ДНК для решения одной из версий “задачи коммивояжера”. Потребовалась всего неделя для получения ответа, в то время как традиционным компьютерам понадобилось бы несколько лет. При этом было использовано фундаментальное явление, свойственное молекулам ДНК - способность к так называемым комплементарным взаимоузнаваниям. Это явление заключается в том, что любые фрагменты каждой из двух цепочек ДНК находят в растворе (или в составе хромосом живой клетки) только собственные, в некотором смысле зеркальные, половинки и образуют нормальную двойную спираль. Успешность и быстрота автоматических поисков половинками ДНК друг друга как акта самоорганизации (самосборки) и обеспечили высокую скорость перебора вариантов в пределах “задачи коммивояжера”. Причины быстрых и точных взаимоузнаваний половинок ДНК до недавнего времени были неизвестны. А это необычайно важно для реального создания ДНК-компьютера, и об этом речь пойдет ниже.
Путь, который выбрал Адлеман, используя ДНК, не то чтобы неверен, скорее, он похож на попытки понять, как, например, происходит процесс мышления у Иванова, Петрова или Сидорова на основе нашего знания о том, что они любят вкусно поесть. Правильное и эффективное использование ДНК, как основного информационного элемента будущего биокомпьютера, немыслимо без понимания истинных функций генетических молекул в биосистемах. Возвращаясь к предыдущим главам, хромосомный аппарат, как система записывающая, сохраняющая, изменяющая и транслирующая информацию, может рассматриваться одновременно на уровнях вещества и достаточно хорошо изученных физических полей, которыми, как носителями генетической и общерегуляторной информации, оперирует континуум генетических молекул (ДНК,РНК). Континуум этот является основным компонентом совокупности хромосом, являющейся, по сути, биокомпьютером. Уровни вещества и поля, на которых хромосомный биокомпьютер функционирует, неразрывны и функционально дополняют друг друга. Здесь реализуются неизвестные ранее виды памяти (солитонная, голографическая, фантомная) и при этом молекулы ДНК могут работать как биолазеры и одновременно как среда записи лазерного сигнала. Кроме того, мы обнаружили, что ДНК способна излучать широкополосное сверхслабое электромагнитное поле, которое нам удалось усилить в тысячи раз. Впрочем, ДНК в этом плане является частным случаем, поскольку зафиксированное нами явление свойственно, вероятно, всем веществам [42], но хромосомы используют этот феномен, наверное, в высшей степени эффективно как один из волновых каналов информационных и (или) энергетических коммуникаций. Молекулы ДНК, как континуум любой биосистемы, способны к формированию прообразов биоструктур и организма в целом как “волновых копий” или “матриц” и сравнению построенного организма с ними как с реперами. В этом плане механизм быстрого и точного взаимоузнавания цепочек (половинок) ДНК, механизм, которым воспользовался Адлеман для решения “задачи коммивояжера”, - лишь один из способов самоорганизации биосистем. Взаимоузнавание, в частности, происходит потому, что в молекулах ДНК зарождаются особые сверхустойчивые акустико-электромагнитные волны (так называемые солитоны), некоторые разновидности которых можно трактовать в рамках открытого в 1949г. “явления возврата Ферми-Паста-Улама” (ФПУ). Такие солитоны ДНК обладают двумя связанными типами памяти - собственно памятью, свойственной явлению ФПУ-возврата, т.е. способностью помнить начальные моды возбуждений и периодически к ним “возвращаться”. Другая память ДНК-континуума в биосистеме - квази-голографическая или фрактальная. Она связана с фундаментальным свойством биосистем - восстанавливать целое из своей части. Это свойство фудаментально и хорошо известно (черенкование растений, регенерация хвоста у ящериц, регенерация целого организма из яицеклетки). Высшая форма такой памяти - ассоциативная память коры головного мозга, т. е. нейронов. Бесперспективно рассуждать о ДНК-компьютере, даже решив с помощью молекул ДНК “задачу коммивояжера”, если не учитывать новую логику в понимании знаковых, кодирующих биофункций ДНК. Другая сторона дела состоит в соотнесении этой логики с многочисленными исследованиями по нейрокомпьтерам и попытками разобраться в “компьютерной” работе мозга без понимания кодирующих функций нервного импульса.
Принято считать, что нервный импульс прост, что он является всего лишь волной деполяризации, и поэтому шифровка информации здесь происходит только по частотному механизму. Расчеты показывают, однако, что частотные модуляции недостаточны для кодирования. А.А.Березиным проведено крупное исследование, из которого следует, что нервные импульсы - это все те же солитоны в рамках явления ФПУ-возврата, при этом, самое главное, такие солитоны отображают в своей структуре РНК-”тексты”, а следовательно, и ДНК В развитии этой идеи нами выдвинуто предположение о знаковой смысловой связи солитонов на хромосомном уровне и солитонов нервных импульсов. Это свидетельствует в пользу связи сознания и его отображения - слова с основной информационной фигурой биосистем - молекулами ДНК с их новыми, неизвестными ранее, типами памяти. Солитоны ДНК способны после “прочитывания ими генотекстов” покидать пределы ДНК в форме особых электромагнитных полей с тем, чтобы узнавать нужные участки других молекул ДНК и формировать целостные новые “тексты”, необходимые в данный момент для выполнения биофункций, в том числе и путем передачи информации в нервные импульсы. Что касается термина “тексты ДНК”, который раньше употреблялся для удобства и был взят взаймы у лингвистов для метафорического употребления, то оказывается, эта текстовая структура ДНК действительно сродни человеческой речи. Наши математико-лингвистические исследования [14,25, см. также предыдущие главы] показали, что такой ключевой параметр, как фрактальность, един для ДНК и человеческой речи. Это коррелирует с ранними работами в этой области
Используя эти теоретические разработки и собственные данные по физико-химии ДНК, нам удалось экспериментально доказать возможность свертки генетической информации в форме солитонных волновых пакетов, описываемых физико-математическим формализмом явления возврата Ферми-Паста-Улама (ФПУ). Такие волновые пакеты с искусственно введенной в них биоинформацией, продуцируемые соответствующими ФПУ-радиоэлектронными устройствами, способны входить в резонансный информационный контакт с генетическим аппаратом животных, растений и, вероятно, человека с последующим резким и направляемым изменением их обмена веществ. Оказалось, что вещество наследственности - ДНК - является генератором ФПУ-солитонных акустических и электромагнитных полей. Именно поэтому ФПУ-генераторы способны вводить информацию в хромосомы по электромагнитным резонансным механизмам. Эффективность генераторов на порядки возрастает, если практически использовать феномен математической общности фрактальной структуры ДНК-”текстов” и человеческой речи. Грамматика генетических текстов является, вероятно, частным случаем универсальных грамматик всех языков людей. Поэтому и реализуются физико-смысловые резонансы солитонных структур ДНК и искусственных ФПУ-солитонных полей, как аналогов естественных ФПУ-хромосомных полей.
Вводя определенные кодовые вербальные команды через генератор ФПУ в генетический аппарат радиационно поврежденных семян растений, удалось достоверно уменьшить число хромосомных аберраций, т.е фактически блокировать поражающее действие рентгеновского облучения. Более того, оказалось, что возможна превентивная защита генома растений от жесткого рентгеновского излучения с помощью адекватных волновых команд. Контрольные эксперименты с хаотическими вербальными построениями (командами), введенными через ФПУ-устройства в геном биосистем, показали, что такие команды никак не влияют на целостность хромосом.
Эти эффекты предсказаны и проверены на основании использования математических компьютерных моделей, имитирующих “чтение” солитонами на ДНК генотекстов и ретрансляцию этих текстов в другие клетки и ткани.
Другие наши физико-математические модели и эксперименты обосновывают так называемый “антенный эффект” при возбуждении электромагнитными полями выделенных коллективных мод макромолекул ДНК. Это прямо связано с экспериментами по так называемому двухфотонной накачке геноструктур с последующим лазерным излучением ДНК [18] и также согласуется с нашими результатами по взаимодействию гелей ДНК с импульсным излучением инфракрасного лазера [25].
Вновь вернемся к компьютеру на ДНК. Ясно, что при его разработке необходимо использовать не только и не столько результаты эксперимента Адлемана. Чтобы реализовать свои возможности, ДНК должна находиться в привычной среде - в водном растворе или в жидкокристаллическом состоянии. Но это лишь начало. Другие возможности ДНК или хромосом могут быть выявлены в условиях, приближенных к тем, которые имеются в живой клетке. В пределе компьютер на ДНК - это и есть живая клетка, то есть надо создать искусственный организм, а это произойдет не скоро. Сейчас мы можем делать только какие-то приближения к состоянию ДНК в клетке, но и это немало. Можно перечислить то, что реально выполнимо уже сейчас. Прежде всего необходимо начать практическое использование новых типов памяти геноструктур и для этого пытаться конструировать ячейки памяти, работающие на явлении ФПУ-резонансов и (или) на способности записывать голограммы. Такая память будет на многие порядки по объему и быстродействию превосходить память существующих магнитных, оптических дисков и голографических систем. Вторая принципиальная возможность связана с этими типами памяти, но многократно усиливается способностью хромосом быть лазеро-активной средой. Препараты хромосом выступают в таком варианте и как ячейка памяти, и как лазеры, считывающие собственную (а также наведенную) голографическую и ФПУ-память. И, наконец, последняя из достижимых в настоящее время - использование квази-речевых характеристик ДНК. Можно создавать такие ДНК-лазеры, которые будут высвечивать и “озвучивать” как естественные генотексты, так и искусственные (синтезированные) знаковые последовательности полинуклеотидов, имитирующие естественные генопрограммы. Однако это весьма опасный путь и необходима система запретов на искусственные волновые гены. Такой способ работы с ДНК-компьютерами означает вхождение в новые семиотические ареалы генома человека, вообще всей биосферы, ареалы, которые Природа (или Бог) использовала для создания человека. Мысль вполне реалистичная, если учесть теоретические работы по коллективной симметрии генетического кода, проводимые школой Эйгена в Институте Макса Планка. Ее исследования показывают, что ключевая часть информации, записанная и записываемая как квазиречь в хромосомах всех организмов нашей планеты, носит искусственный характер. Наши данные о том, что хромосомный континуум и ДНК любой биосистемы является неким подобием антенны, открытой во вне для приема дополнительной (возможно, экзобиологической) информации, подтверждают сказанное. Можно думать, что геном организмов Земли, по крайней мере частично, является полигоном для смысловых экзобиологических влияний, и в этом плане существенно, что мы нашли первичные подходы к вхождению в этот семиотико-семантический ареал.
Основываясь на сказанном, можно предсказать, что открываются следующие перспективы знаковых манипуляций с геноструктурами как основным субстратом биокомпьютеров:
1. Создание искусственной памяти на генетических молекулах, обладающей поистине фантастическим объемом и быстродействием.
2. Создание биокомпьютера на ДНК, основанного на совершенно новых принципах и сравнимого по способам обработки информации и функциональным возможностям с человеческим мозгом.
3. Осуществление дистантного управления ключевыми информационными процессами в биосистемах через искусственные биокомпьютеры (лечение рака, СПИДа, генетических уродств, управление социогенетическими процессами и, в конечном итоге, изменение времени жизни).
Похожие работы
... нравственно и этически взвешенного подхода, поскольку произвольные коррективы здесь могут привести к быстрому саморазрушению человечества и всего Живого на Земле. В связи с развитием идей волновой (и "речевой") генетики необходима система жестко определенных запретов в экспериментах в этой нарождающейся области знания, подобная существующей в генетической инженерии. Идея квазивербального или, что ...
... режима". Поощряемая на государственном уровне "массовая культура" демонстрирует пародирование и кощунство, опошление и оскорбление святынь, в том числе исторического прошлого, издевательство над великой русской культурой. Тому свидетельство — насаждаемая сверху швыдковщина: непечатная брань, демонстративная пошлость, зловещий разлив русофобии и опустошение духовного начала в воспитании школьников ...
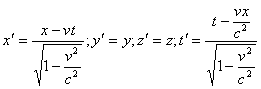
... инерциальных системах отсчета. Пространственно-временной континуум – неразрывная связь пространства и времени и их зависимость от системы отсчета. Тема 11. Основные концепции химии 1. Химия как наука, ее предмет и проблемы Важнейшим разделом современного естествознания является химия. Она играет большую роль в решении наиболее актуальных и перспективных проблем современного общества. К ...
сте с этим закончился прежний, классический этап в развитии естествознания, характерный для эпохи Нового времени. Наступил новый этап неклассического естествознания XX века, характеризующийся, в частности, новыми, квантово-релятивистскими представлениями о физической реальности. 2. Научно-техническая революция и ее естественнонаучная составляющая Новые явления и процессы, имевшие место в ...
0 комментариев