Навигация
Активно защищаться от деструктивных волновых влияний через обнаруженный информационно-энергетический канал
177379
знаков
0
таблиц
29
изображений
4. Активно защищаться от деструктивных волновых влияний через обнаруженный информационно-энергетический канал.
5. Устанавливать экзобиологические контакты.
Подводя итог, спросим : что остается от логики постановочных экспериментов с ДНК, которую предлагают Адлеман и другие исследователи в области молекулярной электроники информационных биомакромолекул? Эта логика уязвима, поскольку основана на упрощенных представлениях о работе хромосом как чисто вещественного субстрата. Волновые функции геноструктур не берутся в расчет. Это тупик, который оборачивается все более нарастающим огромным финансированием по гено-биотехнологиям, по нейрокомпьютерам со все меньшим практическим выходом. Те же пороки ожидают и молекулярную электронику в ее попытках использовать одномерное мышление относительно ДНК при создании биокомпьютера.
Такой компьютер должен имитировать функции генома в части оперирования волновой информацией - то есть создавать образы, в том числе и квази-речевые, распознавать их, манипулировать ими как командными. Такие знаковые структуры будут обладать огромной биологической активностью. Даже сейчас лазер на ДНК, “заряженный” определенными текстами, например, геном продолжительности жизни, вероятно, мог бы продлить ее у человека на 300 - 400 и более лет. Необходимо перераспределение финансирования в генетике, эмбриологии и генной инженерии, а также в молекулярной электронике. Это позволит сделать прорыв в создании компьютеров с квази-генетической памятью, объем которой превосходит все мыслимые пределы, и способных управлять суперсложными процессами, реально сравнимыми с метаболизмом и мышлением. Такие биокомпьютеры будут способны контролировать и поддерживать нормальную жизнедеятельность человека во временных масштабах, приближающихся к бессмертию.
ЯВЛЕНИЕ ПЕРЕХОДА СВЕТА В РАДИОВОЛНЫ ПРИМЕНИТЕЛЬНО К БИОСИСТЕМАМ И БИОКОМПЬЮТЕРАМ [42]
Нами обнаружено явление перехода красного когерентного света в радиоволны при взаимодействии лазерного пучка с веществами. Рассмотрим, как формируются на порядки усиленные радиоизлучения геноструктур in vitro, формируемые за пределами лазерного резонатора, основной частью которых могут являться электроакустические поля, обнаруженные нами ранее, в частности, как волновые структуры ДНК, рибосом, коллагена и хромосом [25].
Для получения стоячей и бегущих волн на лазерном пучке, зондирующем геноструктуры, в наших экспериментах был использован специально изготовленный He-Ne лазер с мощностью излучения 2 mВт, длиной волны 632,8 нм, одночастотный со стабильным резонатором, управляемым посредством термостатирующего элемента. Схема эксперимента приводится на рис 15.
Это схема так называемого трехзеркального лазерного интерферометра имеет особенность в том, что внешнее зеркало резонатора является полупрозрачным или полностью прозрачным, а также может быть преобразовано в сложные композиции типа “сэндвич”, между слоями которого вводятся изучаемые препараты. В этой схеме имеются два информа-
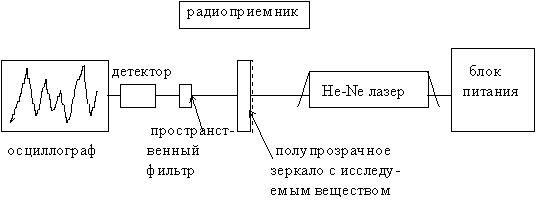 Рис.15
Рис.15
ционных канала - один традиционный оптический, а другой по эфиру - радиоволновый. Рассмотрим вначале работу оптического канала для того, чтобы понять работу второго канала. Внешнее зеркало резонатора юстируется таким образом, чтобы отраженная часть оптического луча от этого зеркала точно попадала бы в резонатор. При этом прямой из лазера и отраженный от зеркала лучи интерферометра складываются и результирующая волна от сложения сигналов, прошедшая через полупрозрачное зеркало, анализируется пространственным фильтром и попадает на кристалл фотодетектора. Пространственный фильтр, представляющий собой точечную диафрагму, жестко соединен в едином моноблоке с корпусом фотодетектора, электрический сигнал от которого наблюдается на экране осциллографа. В зависимости от того, в какой спекл попадает точечный участок фильтра, наблюдаются те или иные осциллографические сигналы, которые появляются за счет резонансного усиления радиоизлучений, формируемых за пределами лазерного резонатора.
Регистрацию радиоволн проводили “радиопрослушиванием” близлежащего пространства (в радиусе до 4-х метров) вокруг препаратов специальным образом приготовленных хромосом (сперматозоиды мыши) и ДНК из селезенки быка. Концентрированные препараты (около 1 мг/мл в дистиллированной воде, а также в определенных сочетаниях с этанолом, водой, триптофаном и гуанозин-трифосфатом) наносили тонким слоем на грань полупрозрачного зеркала (предметное стекло), накрывали вторым предметным стеклом, выдерживали 3 - 4 суток до высыхания при
![]() , и направляли отраженный (промодулированный исследуемым препаратом) пучок света обратно в лазерный резонатор. В другой модификации использовали тонкую, отражающую свет, пленку ДНК без нанесения ее на полупрозрачное зеркало. Схема эксперимента с воздействием ультразвука на ДНК на рис.16.
, и направляли отраженный (промодулированный исследуемым препаратом) пучок света обратно в лазерный резонатор. В другой модификации использовали тонкую, отражающую свет, пленку ДНК без нанесения ее на полупрозрачное зеркало. Схема эксперимента с воздействием ультразвука на ДНК на рис.16.
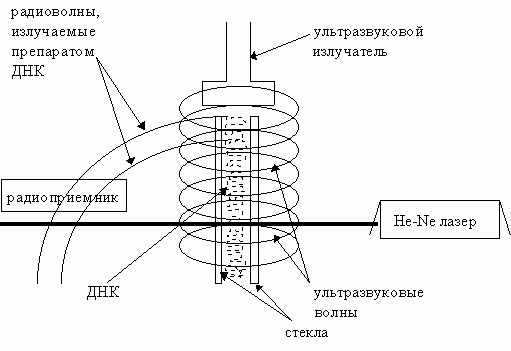
Рис.16
При всех способах подготовки ДНК регистрировали отчетливые радиосигналы, различающиеся по характеру в зависимости от типа исследуемых образцов или их сочетаний. Зондирование ДНК и хромосом и их комплексов с перечисленными выше веществами сопровождалось особыми радиосигналами (радиозвуком), резко отличным от такового абиогенных препаратов. Характерным, и полностью совпадающим с данными [25], было наличие чередования четких одиночных радио звуковых периодических (или почти периодических) сигналов, чередующихся со стохастическими, аналогично наблюдению в [25] относительно характера воздействия инфракрасного лазера
![]() на ДНК. Представляется, что это еще одна демонстрация высокой самоорганизации (солитонообразования) ДНК в рамках явления возврата Ферми-Пасты-Улама, самоорганизации, свойственной генетическим структурам как одно из фундаментальных проявлений памяти наследственности. Иными словами, в настоящем исследовании мы в какой-то степени подтвердили обнаруженную в [25] “солитонную память” ДНК на определенные моды возбуждения препарата, например, на механические вибрации. Характерно, что специфические модуляции радио звука при этом полностью соответствовали изменению во времени двумерных спекл-картин рассеянного препаратами ДНК света. Этот эффект полностью прекращался, если на препараты воздействовали ультразвуком (25 кГц, мощность 6,6
на ДНК. Представляется, что это еще одна демонстрация высокой самоорганизации (солитонообразования) ДНК в рамках явления возврата Ферми-Пасты-Улама, самоорганизации, свойственной генетическим структурам как одно из фундаментальных проявлений памяти наследственности. Иными словами, в настоящем исследовании мы в какой-то степени подтвердили обнаруженную в [25] “солитонную память” ДНК на определенные моды возбуждения препарата, например, на механические вибрации. Характерно, что специфические модуляции радио звука при этом полностью соответствовали изменению во времени двумерных спекл-картин рассеянного препаратами ДНК света. Этот эффект полностью прекращался, если на препараты воздействовали ультразвуком (25 кГц, мощность 6,6
![]() ) в течение 10-15 секунд на расстоянии 1-2 сантиметра. После этого радиозвук становился монотонным и практически не отличался от фонового.
) в течение 10-15 секунд на расстоянии 1-2 сантиметра. После этого радиозвук становился монотонным и практически не отличался от фонового.
Можно высказать рабочую гипотезу о том, какие события происходят при взаимодействии ультразвука и генетических структур:
а) происходят разрывы молекул ДНК;
б) некоторые моды (или обертона) “записываются” на уровне акустики ДНК в рамках солитонного явления возврата Ферми-Паста-Улама и периодически возвращаются, “проигрываются” геноструктурами, подавляя (зашумляя) генознаковую ( в том числе и онко-генознаковую) акустику ДНК;
в) происходят оба перечисленных процесса.
В живом организме при хирургических операциях на раковых опухолях ультразвуковыми скальпелями, разработанных В.И.Лощиловым, это приводит к “стиранию” извращенной генетической информации, даваемой онкогенами, и тем самым к прерыванию метастазов.
Это служит основой для разработки принципиально новых методологий “волновой хирургии” онкологических больных, а если шире, то и “волновой медицины”.
Попытаемся дать первичную физическую модель феномена, когда лазерный свет (при взаимодействии с исследуемыми веществами) генерирует радиоволны. В общем случае всякую излучающую систему можно представить в виде потенциального гармонического осциллятора, находящегося в первоначальном состоянии устойчивого равновесия. В нашем случае, когда внешнее зеркало резонатора неподвижно, система находится в состоянии устойчивого равновесия и не излучает радиоволны. При этом (![]() ), для которого потенциал
), для которого потенциал ![]() минимален. При отклонении зеркала из устойчивого равновесия на некоторое расстояние
минимален. При отклонении зеркала из устойчивого равновесия на некоторое расстояние ![]() потенциал в этой точке может быть разложен в ряд по степеням малых величин
потенциал в этой точке может быть разложен в ряд по степеням малых величин ![]() , а так как в этой точке частная производная
, а так как в этой точке частная производная ![]() = 0, то будем иметь значения для потенциала гармонического осциллятора
= 0, то будем иметь значения для потенциала гармонического осциллятора
![]()
![]() .
.
Запишем волновое уравнение для гармонического осциллятора в виде:
![]() , (1)
, (1)
где ![]() - постоянная планка,
- постоянная планка,
![]() - квазиупругая постоянная,
- квазиупругая постоянная,
![]() , где
, где
![]() - угловая частота колебаний, а
- угловая частота колебаний, а
![]() -потенциал.
-потенциал.
Если 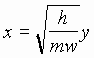 ,
, ![]() , то уравнение (1) примет вид:
, то уравнение (1) примет вид:
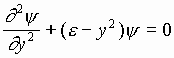 .
.
Известно, что энергия гармонического осциллятора всегда положительна, и для среднего значения энергии ![]() имеем:
имеем:
![]() ,
,
где собственные значения
![]() будут:
будут:
![]() ,
,
и они равны собственным значениям гармонического осциллятора с точностью для полуцелого квантования энергии. В нашем случае имеет место суперпозиция двух (![]() и
и
![]() ) монохроматических гармонических волн от точечных осцилляторов, одним из которых является лазер, а другим внешнее зеркало резонатора. Для упрощения выкладок построения физико-математической модели предположим, что поляризация этих волн одинакова и совпадает по ориентации магнитных
) монохроматических гармонических волн от точечных осцилляторов, одним из которых является лазер, а другим внешнее зеркало резонатора. Для упрощения выкладок построения физико-математической модели предположим, что поляризация этих волн одинакова и совпадает по ориентации магнитных
![]() и электрических
и электрических ![]() векторов Умова-Пойтинга. Результирующее колебание в такой суперпозиции (
векторов Умова-Пойтинга. Результирующее колебание в такой суперпозиции (![]() и
и
![]() ) волн дадут:
) волн дадут:
![]() ,
,
где ![]() и
и ![]() - амплитуды волн;
- амплитуды волн;
![]() и
и ![]() - фазы;
- фазы;
![]() - угловая частота;
- угловая частота; ![]() - время.
- время.
Для волн, распространяющихся в одном направлении, результирующее колебание можно записать как
![]() ,
,
где
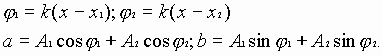 (2)
(2)
Если ![]() , то
, то
![]() .
.
Из уравнения (2), получим новую гармоническую волну, амплитуда и фаза которой будет иметь вид:
![]() ,
,
![]() =
=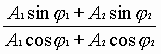 . (3)
. (3)
Угол (![]() ) в выражении для амплитуды результирующей волны равен
) в выражении для амплитуды результирующей волны равен
(![]() )
)![]() .
.
Таким образом, произведя сложение волн посредством третьего зеркала резонатора, которое луч возвращает точно в резонатор лазера, получили новую гармоническую волну. Из последнего выражения видно, что амплитуда не зависит от положения точки, в которой мы исследуем результирующее колебание. Все точки колеблются с одинаковой амплитудой, зависящей лишь от разности расстояний между точками, в которых фазы суммируемых волн равны нулю. Амплитуда будет максимальной, когда разность фаз будет равна ![]()
![]() , где
, где
![]() - целое число. При этом расстояние между внешним зеркалом и резонатором лазера составит
- целое число. При этом расстояние между внешним зеркалом и резонатором лазера составит ![]() , а амплитуда равна
, а амплитуда равна
![]() . В этом случае волны складываются в фазе. Минимальной амплитуда будет при условии, если
. В этом случае волны складываются в фазе. Минимальной амплитуда будет при условии, если
![]()
![]() , а разность путей двух волн
, а разность путей двух волн ![]() . В этом случае амплитуда равна
. В этом случае амплитуда равна ![]() , что соответствует сложению противофазных волн. Если амплитуды общих волн будут равны, т.е.
, что соответствует сложению противофазных волн. Если амплитуды общих волн будут равны, т.е.
![]() , то наименьшая амплитуда будет равна нулю, а максимальная - удвоенной амплитуде суммируемых волн. При равенстве амплитуд фаза результирующего колебания будет равна
, то наименьшая амплитуда будет равна нулю, а максимальная - удвоенной амплитуде суммируемых волн. При равенстве амплитуд фаза результирующего колебания будет равна
![]() и не будет зависеть от положения точки наблюдения. Во всех точках пространства, где происходит интерференция, фаза колебания одинакова, что соответствует стоячим волнам.
и не будет зависеть от положения точки наблюдения. Во всех точках пространства, где происходит интерференция, фаза колебания одинакова, что соответствует стоячим волнам.
Амплитуда этих волн из (3)
![]() . В случае встречных волн с различными амплитудами
. В случае встречных волн с различными амплитудами ![]() . При наложении таких волн образуется стоячая волна с амплитудой
. При наложении таких волн образуется стоячая волна с амплитудой ![]() , а также бегущие волны с амплитудой
, а также бегущие волны с амплитудой ![]() , которые распространяются в направлении волны с большей амплитудой
, которые распространяются в направлении волны с большей амплитудой ![]() . Эти бегущие волны при записи создают фон, над уровнем которого наблюдается модуляция, обусловленная “дышащей” стоячей волной типа трубчато-цилиндрического солитона-бризера. При неподвижном зеркале внешнего резонатора в нашем случае образуется трубчато-цилиндрический квантовый солитон-бризер, стоящий на месте, но “дышащий”.
. Эти бегущие волны при записи создают фон, над уровнем которого наблюдается модуляция, обусловленная “дышащей” стоячей волной типа трубчато-цилиндрического солитона-бризера. При неподвижном зеркале внешнего резонатора в нашем случае образуется трубчато-цилиндрический квантовый солитон-бризер, стоящий на месте, но “дышащий”.
В нелинейной физике такого рода солитоны названы бризерами именно потому, что они, оставаясь на месте или смещаясь вблизи неоднородностей (например, зеркала), “дышат” (от английского breath). Именно это низкочастотное “оптическое солитонное дыхание”, как нам представляется, и генерирует радиоволны ![]() и
и ![]() . “Солитонное акустико-электромагнитное дыхание” свойственно и молекулам ДНК, рибосомам и коллагену в рамках явления возврата Ферми-Паста-Улама. [25]. Колебания жидкокристаллической решетки молекул ДНК приводят к возникновению поперечных волн интенсивности, проявляющихся в изменении диаметра трубчато-цилиндрического квантового солитона-бризера, который может двигаться, порождая широкий спектр радиоволн. Диаметр этого солитона может уменьшаться или увеличиваться при прямом или обратном движении внешнего зеркала резонатора. Систему подвижных колец (проекций трубок), в той или иной степени искаженных (промо-дулированных) исследуемым препаратом, можно наблюдать на экране, установленном между внешним зеркалом резонатора и пространственным фильтром фотодетектора. Движение такого солитона-бризера может привести к усилению тех бегущих волн интенсивности, о которых речь шла выше. Таким образом, происходит взаимная энергетическая подпитка бегущих волн солитоном-бризером и наоборот. Можно полагать, что подкачка расходуемой энергии солитона-бризера происходит из фоновой стохастической энергии kT за счет стохастического резонанса через фильтр согласования между кратными гармониками макро- и микросистем, где макросистема есть солитон-бризер, а микросистемой является отдельный квантовый осциллятор, например, молекула ДНК.
. “Солитонное акустико-электромагнитное дыхание” свойственно и молекулам ДНК, рибосомам и коллагену в рамках явления возврата Ферми-Паста-Улама. [25]. Колебания жидкокристаллической решетки молекул ДНК приводят к возникновению поперечных волн интенсивности, проявляющихся в изменении диаметра трубчато-цилиндрического квантового солитона-бризера, который может двигаться, порождая широкий спектр радиоволн. Диаметр этого солитона может уменьшаться или увеличиваться при прямом или обратном движении внешнего зеркала резонатора. Систему подвижных колец (проекций трубок), в той или иной степени искаженных (промо-дулированных) исследуемым препаратом, можно наблюдать на экране, установленном между внешним зеркалом резонатора и пространственным фильтром фотодетектора. Движение такого солитона-бризера может привести к усилению тех бегущих волн интенсивности, о которых речь шла выше. Таким образом, происходит взаимная энергетическая подпитка бегущих волн солитоном-бризером и наоборот. Можно полагать, что подкачка расходуемой энергии солитона-бризера происходит из фоновой стохастической энергии kT за счет стохастического резонанса через фильтр согласования между кратными гармониками макро- и микросистем, где макросистема есть солитон-бризер, а микросистемой является отдельный квантовый осциллятор, например, молекула ДНК.
Первоначальная идея использования энергии kT для подпитки солитонов обсуждена в работе Пиппарда и в наших последующих исследованиях относительно создания лазеров на информационных биомакромолекулах, в частности, лазеров с использованием солитонных волн [2,3,16,34,35]. Фильтр согласования мы понимаем как инструмент реализации общего принципа гармонического сочетания, то есть непротиворечивого согласованного взаимодействия. Вычислим диапазон спектра частот генерации такого опто-радиоволнового “дыхания” в наших экспериментах за счет движения внешнего зеркала резонатора. Спектр частот резонатора определяется временем прохождения излучением всего тракта от торца дальнего зеркала лазера до внешнего зеркала резона-
тора по известному соотношению:
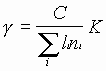 ,
,
где ![]() - целое число;
- целое число;
![]() - длина и показатель преломления среды
- длина и показатель преломления среды
![]() -го участка резонатора;
-го участка резонатора;
![]() - длина резонатора;
- длина резонатора;
![]() -скорость света в среде.
-скорость света в среде.
В резонаторе, как правило, существуют несколько типов бегущих волн, для которых число
![]() различается на единицу и зависит от количества мод. Такой режим является многомодовым. Разность частот между соседними модами составляет
различается на единицу и зависит от количества мод. Такой режим является многомодовым. Разность частот между соседними модами составляет
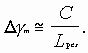
В резонаторе резко сужается ширина спектра излучения. Известно, что добротность резонатора определяется соотношением
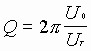 ,
,
где ![]() - энергия, запасенная в резонаторе;
- энергия, запасенная в резонаторе;
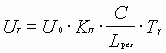 - это энергия, теряемая в резонаторе за цикл времени одного прохода резонатора
- это энергия, теряемая в резонаторе за цикл времени одного прохода резонатора ![]() ,
,
где ![]() -коэффициент суммарных потерь;
-коэффициент суммарных потерь;
![]() -потери энергии за один проход в резонаторе;
-потери энергии за один проход в резонаторе;
![]() - время прохода резонатора.
- время прохода резонатора.
Подставляя значение
![]() в
в ![]() и учитывая, что
и учитывая, что ![]() , получаем:
, получаем:
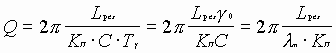 ,
,
так как
![]() , то
, то
![]() ; перепишем
; перепишем
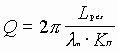 ;
;
Исходя из величины добротности резонатора, находим ширину спектра резонатора:
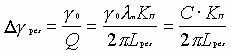 , (4)
, (4)
который является диапазоном спектра излучения радиоволн.
Из работы известна связь между шириной спектра резонатора и шириной линии излучения:
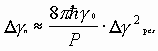 , (5)
, (5)
где ![]() - мощность излучения на данной моде;
- мощность излучения на данной моде;
![]() - энергия кванта.
- энергия кванта.
Простой расчет из формулы (4) показывает, что при
![]() =10%;
=10%; ![]() = 0,54м;
= 0,54м;![]() = 632,8нм;
= 632,8нм;
![]() 1мВт
1мВт
![]() и из формулы (5)
и из формулы (5)
![]() = 0,007Гц.
= 0,007Гц.
Это трансформированные из лазерного света радиоволны в мегагерцовой области, что и было зарегистрировано в наших экспериментах.
Детальное развитие нашей квантово-электродинамической модели порождения радиоволн из света связано с теорией поля Максвелла и идеей возникновения элементарных частиц, которые обладают ротационной осцилляцией электромагнитных векторов Умова-Пойтинга. Эта модель в дальнейшем позволит углубить понимание механизма перехода от пространства к веществу и наоборот и далее к пересмотру понятия времени. Практическое приложение данной теории позволит объяснить такие “аномальные” и давно известные эффекты, как сверхустойчивость термофильных бактерий к температурам, существенно превышающим области термодинамической устойчивости информационных биополимеров - ДНК и белков, а также “аномально” высокую сопротивляемость некоторых биосистем к огромным уровням радиоактивности. В свою очередь, это может вывести на понимание принципов старения и сопротивления старению организмов через идею согласующих фильтров биосистем и окружающей среды, фильтров, фрактально повторяющих себя на микро- и макромасштабах (от элементарных частицы до бактерий и человека).
Имеется косвенное теоретическое подтверждение нашей модели генерации радиоволн из солитонных возбуждений. В работе Тужински и соавторов показана связь, взаимодополняемость двух казалось бы независимых теорий, в которых рассматриваются две физические модели, объясняющие необычное поведение биологических систем. Модели предложены Гербертом Фрелихом и Александром Давыдовым. Так называемые Давыдовские солитоны, описывающие возбуждение, делокализацию и движение электронов вдоль пептидных цепей белковых молекул в форме уединенных волн (солитонов), дополняют известную модель Фрелиха, развитую в нашей работе [16] о возможности высокополяризованного (когерентного, лазероподобного) состояния колеблющихся диполей информационных биомакромолекул, диполей, возникающих при Бозе-конденсации фононов электромагнитных волн белков (![]() ), ДНК (
), ДНК (![]() ), мембран (
), мембран (![]() ). В работе [см. сноску 2 на стр. 85] Давыдовский гамильтониан трансформирован в нормальные координаты, Фрелиховский гамильтониан канонически трансформирован в эквивалентную форму в рамках аппроксимации Хартри-Фока. Авторы полагают, что модель гамильтониана способна связать обе теории, которые математически эквивалентны.
). В работе [см. сноску 2 на стр. 85] Давыдовский гамильтониан трансформирован в нормальные координаты, Фрелиховский гамильтониан канонически трансформирован в эквивалентную форму в рамках аппроксимации Хартри-Фока. Авторы полагают, что модель гамильтониана способна связать обе теории, которые математически эквивалентны.
Кроме того, обе модели дополняют друг друга физически. Бозе-конденсация вибрационных мод биополимеров соответствует распространению солитона волны поляризации. И наоборот, солитонный транспорт граничной энергии вдоль пептидной цепи сопровождается бозе-конденсацией решеточных вибраций биоструктур.
Отсюда следует, что солитон порождает электромагнитное поле, а это, возможно, тот самый эффект, который мы наблюдаем в экспериментах, когда осциллирующий оптический солитон-бризер, отображающий солитонные возбуждения ДНК, генерирует оптико-резонансно усиленные радиоволны.
Еще одна мысль, привлекающая внимание: конверсия эндогенных когерентных фотонов, генерируемых хромосомами, в радиоволны в биосистеме может происходить по трехзеркальному принципу на многочисленных отражающих поверхностях мембран, аналогично нашим модельным опытам. Именно этим может объясняться зеркальный блеск органов и тканей живых организмов. Это же дает надежду, что мы сумеем in vitro-in vivo манипулировать световыми лазерными потоками, которые проводятся сложнейшей сетью световодов живой клетки и которые преобразуются на биомембранах в знаковые радиоволны. Такие процессы могут использоваться как основа для создания принципиально иных компьютеров, точнее биокомпьютеров.
ОСНОВНЫЕ РЕЗУЛЬТАТЫ
1. Проведено математическое (компьютерное) моделирование солитонных возбуждений в молекуле ДНК, показавшее способность уединенных волн (на уровне крутильных колебаний нуклеотидов) реагировать на первичную структуру ДНК и тем самым служить потенциальной основой для истантной трансляции текстовой и иной образной управляющей информации генома.
2. Экспериментально зафиксированы солитоноподобные процессы в гелях ДНК in vitro как реализация явления возврата Ферми-Паста-Улама (ФПУ). Тем самым продемонстрирована принципиальная возможность функционирования генетического аппарата биосистем с использованием ФПУ-волнового типа памяти генома и, соответственно, неизвестного ранее способа передачи геноуправляющих сигналов in vivo полевым путем.
3. Экспериментально доказана возможность существования эпигенетической информации, конвертированной в форму широкополосного электромагнитного поля, в котором реализуется пространственно-временной возврат ФПУ. Доказано также, что такая информация может искусственным способом с помощью генератора ФПУ передаваться от биосистемы-донора к биосистеме-акцептору, что соответствует, вероятно, процессам волнового “метаболизма” эпигеносигналов in vivo. Такой способ является, вероятно, примитивной имитацией главного информационного канала, связанного с Творцом.
4. Найдена компьютерная модель фрактального представления, которая показала, что человеческая речь (тексты) и последовательности нуклеотидов (тексты ДНК) обладают близкой математической структурой.
5. Управление развитием высших биосистем происходит с использованием материально-волновых солитонно-голографических матриц генома, а также семиотических единиц, сходных с человеческой речью и, вероятно, с речью Творца.
6. Разработаны технические устройства и с их помощью найдены первичные основы волнового управления биосистемами на основе явления возврата Ферми-Паста-Улама и эпигено-лингвистических функций генетических молекул.
ЛИТЕРАТУРА
1. Anderson S. et al. Sequence and organization of the human mitochondrial genome. // Nature.1981. v. 290. p. 457- 465.
2. Berezin A.A., Gariaev P.P., Gorelik V.S., Reshetniak S.A., Shcheglov V.A. Is it possible to create laser on information biomacromolecules? // Laser Physics.1996. v.6. № 6. p.1211- 1213.
3. Berezin A.A., Gariaev P.P., Reshetniak S.A., Shaitan K.V., Shcheglov V.A. To the problem of possible development at a biolaser working on Frolich modes. // препринт Физического Института им. П.Н.Лебедева. 1996. № 49. с.12.
4. Crick F.H.C. Codon-anticodon pairing: the wobble hypothesis.// J. Mol. Biol. 1966.v. 19.p.548- 555.
5. Gariaev P.P. “In vitro-in vivo” DNA conjugation with brain activity and the supreme intellect. // Creation Recearch Society Quarterly. (in pr. )
6. Gariaev P.P. DNA as source of new kind of God “knowledge”. Act and Facts.// Impact Series.1994. № 12.p. 7- 11.
7. Gariaev P.P., Acupuncture points as a resonance structures. Act and Facts.// Impact Series. (in pr.)
8. GariaevP.P., Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A. Holographic associative memory of biological systems. SPIE - The International Society for Optical Engineering. CIS Selected Papers. // Coherent Measuring and Data Processing Methods and Devices.1991.v.1621.p.280- 291.
9. Kirkwood T.B., Rosenberg R.F., Galas D.J., eds. Accuracy in molecular Processes: Its control and Relevance to living Systems.// Chapmen and Hall. London, 1986. (Chapters 4- 6,11).
10 Kuo-Chen Chou. Low-frequency collective motion in biomacro-molecules and its biological functions. // Biophys. Chem.. 1988. v. 30. №1. p. 3- 48.
11. Lagerkvist U. “Two out of Three”: an alternative method for codon reading.Proc.// Natl. Acad. Sci. USA., 1978.v. 75. p. 1759- 1762.
12. Mantegna R.N., Buldyrev S.V., Goldberger A.L., Havlin S., Peng S.-K., Simons M. and Stanley H.E. Linguistic Features of Noncoding DNA Sequences. // Phys. Rev. Lett. 1994.v.73. № 23. p.3169- 31
Похожие работы
... нравственно и этически взвешенного подхода, поскольку произвольные коррективы здесь могут привести к быстрому саморазрушению человечества и всего Живого на Земле. В связи с развитием идей волновой (и "речевой") генетики необходима система жестко определенных запретов в экспериментах в этой нарождающейся области знания, подобная существующей в генетической инженерии. Идея квазивербального или, что ...
... режима". Поощряемая на государственном уровне "массовая культура" демонстрирует пародирование и кощунство, опошление и оскорбление святынь, в том числе исторического прошлого, издевательство над великой русской культурой. Тому свидетельство — насаждаемая сверху швыдковщина: непечатная брань, демонстративная пошлость, зловещий разлив русофобии и опустошение духовного начала в воспитании школьников ...
... инерциальных системах отсчета. Пространственно-временной континуум – неразрывная связь пространства и времени и их зависимость от системы отсчета. Тема 11. Основные концепции химии 1. Химия как наука, ее предмет и проблемы Важнейшим разделом современного естествознания является химия. Она играет большую роль в решении наиболее актуальных и перспективных проблем современного общества. К ...
сте с этим закончился прежний, классический этап в развитии естествознания, характерный для эпохи Нового времени. Наступил новый этап неклассического естествознания XX века, характеризующийся, в частности, новыми, квантово-релятивистскими представлениями о физической реальности. 2. Научно-техническая революция и ее естественнонаучная составляющая Новые явления и процессы, имевшие место в ...
0 комментариев