Навигация
РАЗНООБРАЗИЕ ВЕКТОРНЫХ МОЛЕКУЛ
43921
знак
0
таблиц
5
изображений
1.2.4. РАЗНООБРАЗИЕ ВЕКТОРНЫХ МОЛЕКУЛ
Под понятием "вектор" понимается молекула нуклеиновой кислоты, способная после введения в клетку к автономному существованию за счет наличия в ней сигналов репликации и транскрипции.
Векторные молекулы должны обладать следующими свойствами:
1) способностью автономно реплицироваться в клстке-реципиенте, то есть быть самостоятельным репликоном;
2) содержать один или несколько маркерных генов, благодаря экспрессии которых у клетки-реципиента появляются новые признаки, позволяющие отличить трансформированные клетки от исходных;
3) содержать по одному или, самое большее, по два участка (сайта) для различных рестриктаз в разных районах (в том числе в составе маркерных генов), но не в области, ответственной за их репликацию.
В зависимости от целей эксперимента векторы можно условно разделить на две группы: 1) используемые для клонирования и амплификации нужного гена; 2) специализированные, применяемые для экспрессии встроенных чужеродных генов. Вторая группа векторов объединяет векторы, призванные обеспечить синтез белковых продуктов клонированных генов. Векторы для экспрессии содержат последовательности ДНК, которые необходимы для транскрипции клонированных копий генов и трансляции их мРНК в штаммах клеток.
В качестве прокариотических векторов используются плазмиды, бактериофаги; в качестве эукариотических векторов применяют вирусы животных и растений, векторы на основе 2 мкм дрожжей и митохондрий и ряд искусственно сконструированных векторов, способных реплицироваться как в бактериальных, так и в эукариотических клетках (челночные векторы).
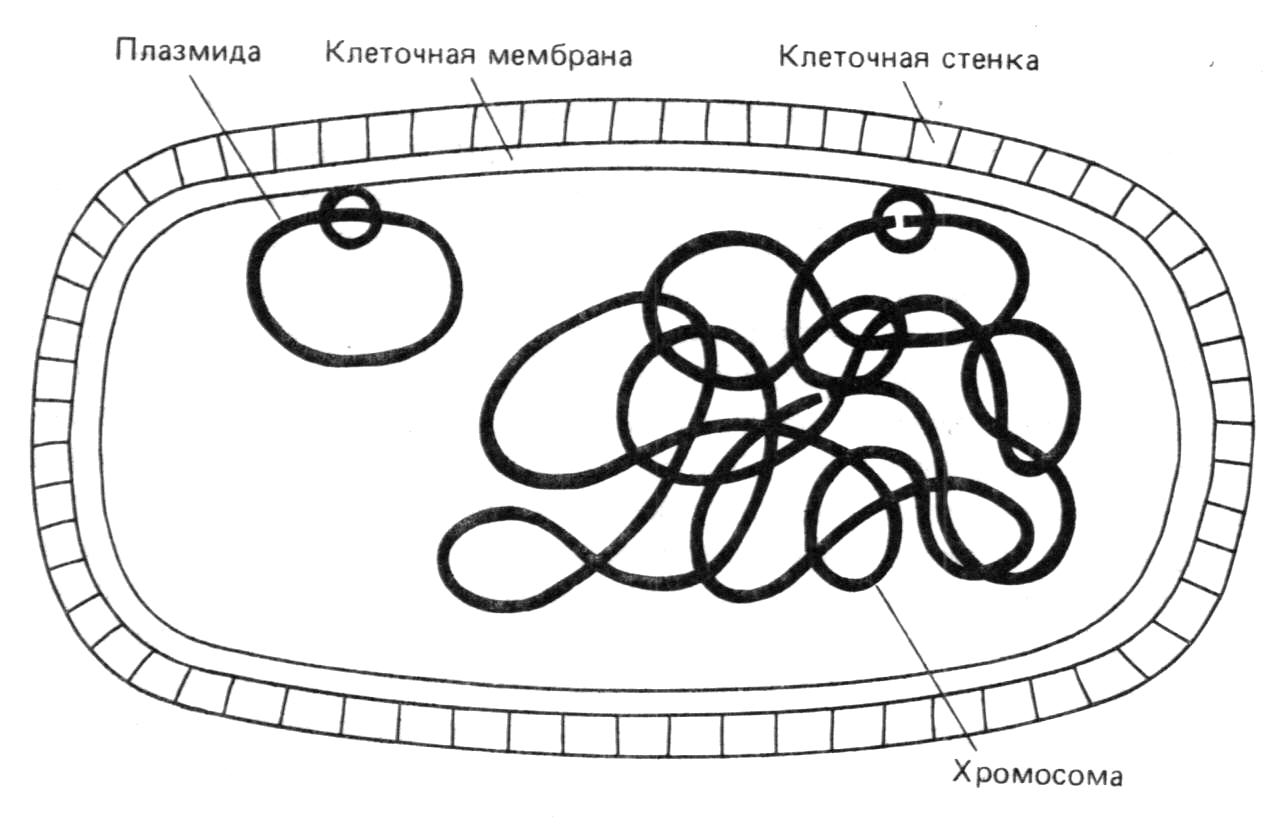
Плазмиды - это внехромосомные генетические элементы про- и эукариот, которые автономно реплицируются в клетках. Большинство плазмидных векторов получено на основе природных плазмид ColE1, pMB1 и p15A.
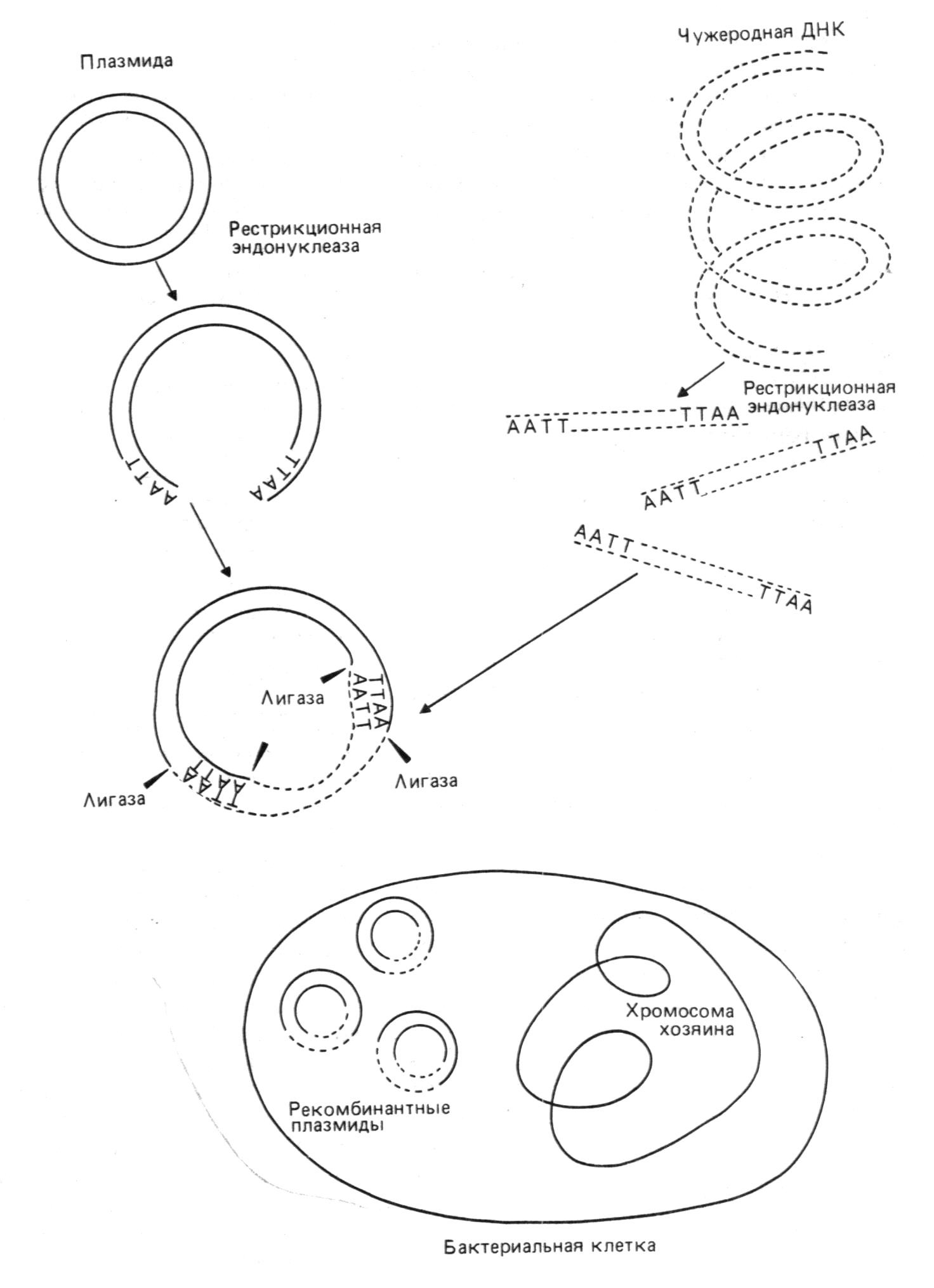
Бактериальные плазмиды делят на два класса. Одни плазмиды (например, хорошо изученный фактор F, определяющий пол у E.coli) сами способны переходить из клетки в клетку, другие такой способностью не обладают. По ряду причин, и прежде всего для предотвращения неконтролируемого распространения потенциально опасного генетического материала, подавляющее большинство бактериальных плазмидных векторов создано на базе плазмид второго класса. Многие природные плазмиды уже содержат гены, определяющие устойчивость клеток к антибиотикам (продукты этих генов - ферменты, модифицирующие или расщепляющие антибиотические вещества). Кроме того, в эти плазмиды при конструировании векторов вводятся дополнительные гены, определяющие устойчивость к другим антибиотикам.
На рис. 4 показан один из самых распространенных плазмидных векторов E.coli - pBR322. Он сконструирован на базе детально изученной плазмиды E.coli - колициногенного фактора ColE1 - и содержит ориджин репликации этой плазмиды. Особенность плазмиды ColE1 (и pBR322 соответственно) состоит в том, что в присутствии ингибитора синтеза белка антибиотика хлорамфеникола (опосредованно ингибирующего репликацию хозяйской хромосомы) ее число в E.coli возрастает от 20-50 до 1000 молекул на клетку, что позволяет получать большие количества клонируемого гена. При конструировании вектора pBR322 из исходных плазмид был делегирован целый ряд "лишних" сайтов для рестриктаз.
В настоящее время наряду с множеством удобных векторных систем для E.coli сконструированы плазмидные векторы для ряда других грамотрицательных бактерий (в том числе таких промышленно важных, как Pseudomonas, Rhizobium и Azotobacter), грамположительных бактерий (Bacillus), низших грибов (дрожжи) и растений.
Плазмидные векторы удобны для клонирования относительно небольших фрагментов (до 10 тыс. пар оснований) геномов небольших размеров. Если же требуется получить клонотеку (или библиотеку) генов высших растений и животных, общая длина генома которых достигает огромных размеров, то обычные плазмидные векторы для этих целей непригодны. Проблему создания библиотек генов для высших эукариот удалось решить с использованием в качестве клонирующих векторов производных бактериофага l.
Среди фаговых векторов наиболее удобные системы были созданы на базе геномов бактериофагов l и М13 E.coli. ДНК этих фагов содержит протяженные области, которые можно делегировать или заменить на чужеродную ДНК, не затрагивая их способности реплицироваться в клетках E.coli. При конструировании семейства векторов на базе ДНК l фага из нее сначала (путем делений коротких участков ДНК) были удалены многие сайты рестрикции из области, не существенной для репликации ДНК, и оставлены такие сайты в области, предназначенной для встраивания чужеродной ДНК. В эту же область часто встраивают маркерные гены, позволяющие отличить рекомбинантную ДНК от исходного вектора. Такие векторы широко используются для получения "библиотек генов". Размеры замещаемого фрагмента фаговой ДНК и соответственно встраиваемого участка чужеродной ДНК ограничены 15-17 тыс. нуклеотидных остатков, так как рекомбинантный фаго -
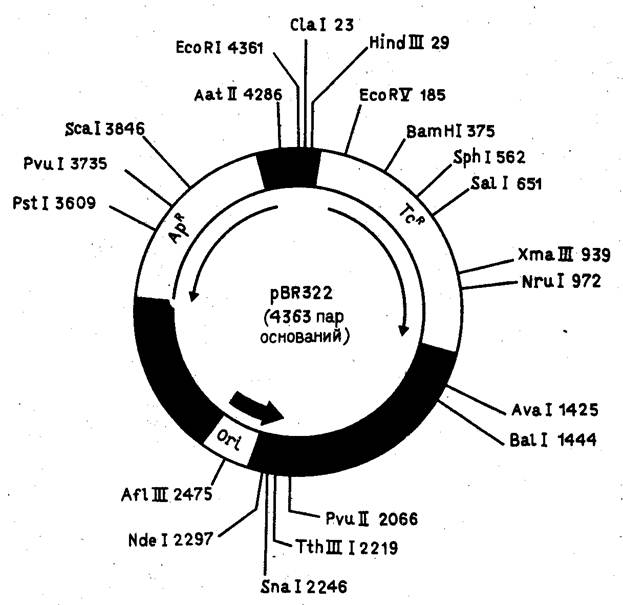
Рисунок 4. Детальная рестрикционная карта плазмиды pBR322.
вый геном, который на 10% больше или на 75% меньше генома дикого l фага, уже не может быть упакован в фаговые частицы.
Таких ограничений теоретически не существует для векторов, сконструированных на базе нитчатого бактериофага М13. Описаны случаи, когда в геном этого фага была встроена чужеродная ДНК длиной около 40 тыс. нуклеотидных остатков. Известно, однако, что фаг М13 становится нестабильным, когда длина чужеродной ДНК превышает 5 тыс. нуклеотидных остатков. Фактически же векторы, полученные из ДНК фага М13, используются главным образом для секвенирования и мутагенеза генов, и размеры встраиваемых в них фрагментов намного меньше.
Эти векторы конструируются из реплекативной (двутяжевой) формы ДНК фага М13, в которую встроены "полилинкерные" участки (пример такой конструкции показан на рис. 5). В фаговую частицу ДНК включается в виде однотяжевой молекулы. Таким образом, этот вектор позволяет получать клонированный ген или его фрагмент как в двутяжевой, так и в однотяжевой форме. Однотяжевые формы рекомбинантных ДНК широко используются в настоящее время при определении нуклеотидной последовательности ДНК методом Сэнгера и для олигодезоксинуклеотид-направленного мутагенеза генов.
Перенос чужеродных генов в клетки животных осуществляется с помощью векторов, полученных из ДНК ряда хорошо изученных вирусов животных - SV40, некоторых аденовирусов, вируса папиломы быка, вируса оспы и так далее. Конструирование этих векторов проводится по стандартной схеме: удаление "лишних" сайтов для рестриктаз, введение маркерных генов в области ДНК, не существенные для ее репликации (например, гена тимидин-киназы (tk) из HSV (вируса герпеса)), введение регуляторных районов, повышающих уровень экспрессии генов.
Удобными оказались так называемые "челночные векторы", способные реплицироваться как в клетках животных, так и в клетках бактерий. Их получают, сшивая друг с другом большие сегменты векторов животных и бактерий (например, SV40 и pBR322) так, чтобы районы, ответственные за репликацию ДНК, остались незатронутыми. Это позволяет проводить основные операции по конструированию вектора в бактериальной клетке (что технически намного проще), а затем полученную рекомбинантную ДНК использовать для клонирования генов в животной клетке.
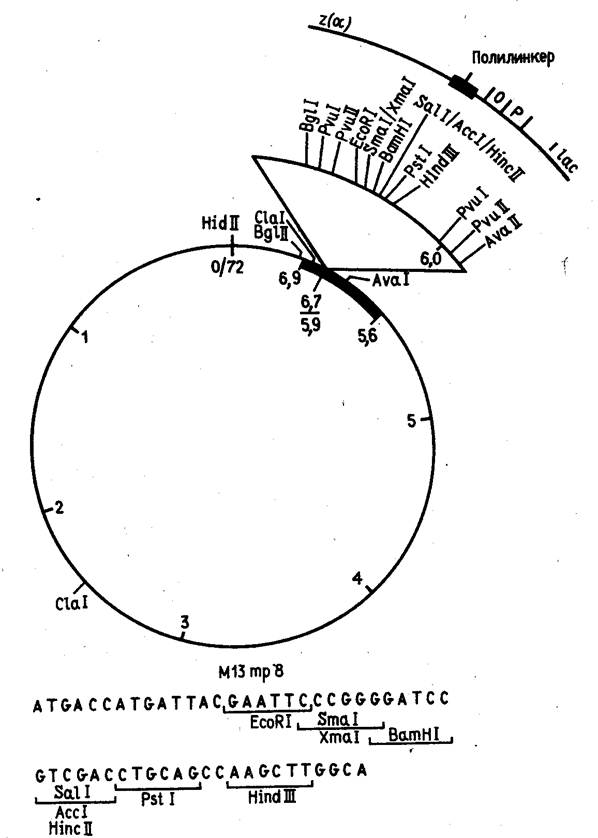
Рисунок 5. Рестрикционная карта вектора М13 mp8.
1.3. ВЫБОР СИСТЕМЫ ЭКСПРЕССИИ КЛОНИРОВАННОГО
ГЕНА, СПОСОБНОЙ ОБЕСПЕЧИТЬ МАКСИМАЛЬНЫЙ ВЫХОД И
ФУНКЦИОНАЛЬНУЮ ПОЛНОЦЕННОСТЬ ПРОДУКТА
Полученные рекомбинантные молекулы переносятся в определенные группы клеток, которые должны обеспечить экспрессию этих генов, то есть синтез соответствующих белков в количествах, экономически рентабельных по сравнению с обычной технологией их производства.
Обычно для данной цели используют бактериальные или дрожжевые культуры клеток, а также системы экспрессии на основе эукариотических клеток.
Из бактериальных клеток наиболее изученной в молекулярно-генетическом отношении является грамотрицательная бактерия Escherichia coli, поэтому для нее можно с наибольшей определенностью планировать генноинженерные конструкции. Однако E. coli слабо освоена промышленностью. Кроме того, она относится к условно-патогенным для человека микроорганизмам, что может создать трудности при получении на ее основе фармацевтических препаратов.
Отмеченные недостатки E. coli легко преодолеваются при конструировании методами генной инженерии штаммов-продуцентов на основе клеток Bacillus subtilis. Данная почвенная бактерия безопасна для человека и животных и прекрасно освоена микробиологической промышленностью. Бактерия B. subtilis по степени изученности следует за E. coli. Важное отличие ее от E. coli - способность эффективно секретировать во внешнюю среду целый ряд белков, поэтому особенно интересны работы по созданию штаммов-продуцентов B. subtilis, секретирующих чужеродные белки из клеток. Однако данная бактерия имеет свои недостатки: рекомбинантные плазмиды в B. subtilis характеризуются нестабильностью, выражающейся в перестройках и делециях ДНК; бациллы секретируют в культуральную среду большое количество протеаз, что существенно усложняет вопрос максимализации генноинженерного получения целевого белка на основе бацилл-продуцентов.
Среди эукариотических микроорганизмов наиболее изученным является низший эукариот Saccharomyces cerevisiae. Одно из преимуществ S. cerevisiae как экспериментальной системы - простота и надежность ее генетического анализа. В клетках дрожжей имеется ферментативная система гликозилирования белков, которая обеспечивает возможность синтеза в них полноценных белков высших эукариот. Аналогичных систем процессинга белков в бактериальных клетках нет. Многие штаммы дрожжей освоены микробиологической промышленностью. Доказана их безвредность для человека и животных. Именно на S. cerevisiae создан первый штамм-продуцент поверхностного антигена вируса гепатита Б, позволивший получить и испытать вакцину против данного вирусного заболевания человека.
С появлением генной инженерии внимание многих исследователей привлекла система культивируемых клеток животных. Особый интерес к культурам клеток животных стал проявляться после обнаружения того, что часть эукариотических генов раздроблена и лишь в системе клеток высших эукариот можно достичь правильной экспрессии таких генов. Кроме того, многие белки животных и их вирусов синтезируются первоначально в виде более высокомолекулярных предшественников, которые в результате специфического протеолитического процессинга переходят в так называемую зрелую форму. Такой процессинг данных белков, по-видимому, можно ожидать лишь в системе клеток животных. Все это убедительно доказывает важность разработки экспрессирующей системы на основе клеток животных.
Для обеспечения наиболее эффективной экспрессии клонированных генов в векторные молекулы встраивают определенные фрагменты ДНК, позволяющие увеличить выход чужеродного белка. Так, для достижения более высокого уровня экспрессии гена HBsAg в клетках E. coli были использованы различные по силе промоторы (промоторы генов cat, kan, bla, trp и тандемно расположенных промоторов генов kan и trp). Уровни синтеза последовательностей HВsAg (нативного и в составе химерных белков) составляли в зависимости от используемых конструкций векторов от 100 до 100000 молекул на клетку.
Похожие работы
... олигонуклеотидов—одну полуавтоматическую, а вторую в комплексе с компьютером. В 1982 г. цена этих приборов на американском рынке составляла 36000—39500 долл.[2]. К открытиям связанным с достижениями генной инженерии нужно прибавить то, что огромный генетический «чертеж» многоклеточного существа просчитан полностью. Я думаю это можно назвать достижением века. После восьми лет работы многих ...
... генно-инженерных исследований. Многие из этих вопросов были подняты самими учеными активно работающих в данной области. В настоящее время большинство исследователей считали, что опасения касающиеся, генной инженерии, не имеют достаточно оснований, но многие этические проблемы остаются нерешенными и продолжают возникать новые. В прошлом генетика и медицинская генетика развивалась как относительно ...

... при постановке биологических или медицинских экспериментов. Составлять его должны ведущие специалисты в этой области и знающие предмет юристы. 3. Другие применения генной инженерии Несмотря на успехи ученых в генной инженерии в таких отраслях, как создание генно-модифицированных продуктов и клонировании животных и человека, они на этом не останавливаются. В целях сохранения военного ...
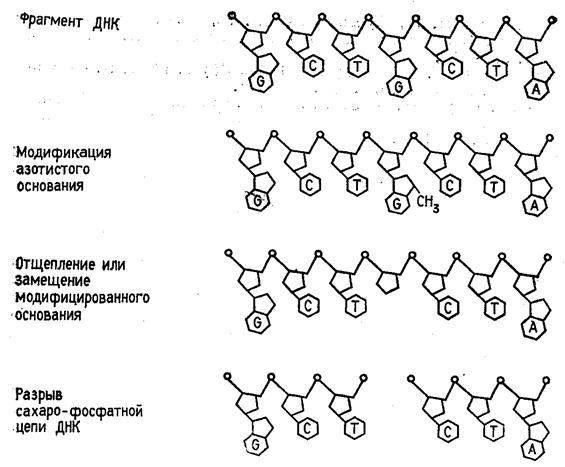
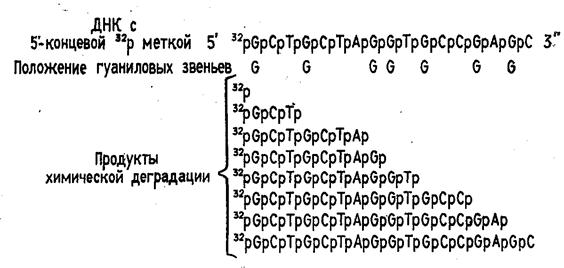
... (например, семейства генов) необходимо провести клонирование ПЦР-продуктов в векторах типа М13, в результате каждый фаг будет содержать только одну вставку. При прямом секвенировании смеси генов наблюдается несколько одинаково расположенных полос в разных дорожках геля. При амплификации же одного гена можно проводить прямое секвенирование, не прибегая к промежуточному субклонированию. Выбор ...
0 комментариев