Навигация
ВЫБОР ВЫСОКОАКТИВНОЙ И ХОРОШО ИЗУЧЕННОЙ В
43921
знак
0
таблиц
5
изображений
1.2. ВЫБОР ВЫСОКОАКТИВНОЙ И ХОРОШО ИЗУЧЕННОЙ В
ИММУНОЛОГИЧЕСКОМ ОТНОШЕНИИ МОДЕЛИ ВЕКТОРА-НОСИТЕЛЯ И КЛОНИРОВАНИЕ СООТВЕТСТВУЮЩЕГО ГЕНА
1.2.1. ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ ДНК
Суть конструирования рекомбинантных ДНК заключается во встраивании фрагментов ДНК, среди которых находится интересующий нас участок ДНК, в так называемые векторные молекулы ДНК (или просто векторы) - плазмидные или вирусные ДНК, которые могут быть перенесены в клетки про- или эукариот и там автономно репли-цироваться. На следующем этапе проводится отбор тех клеток, которые несут в себе рекомбинантные ДНК (с помощью маркерных признаков, которыми обладает сам вектор), и затем индивидуальных клонов с интересующим нас сегментом ДНК (используя признаки или пробы, специфичные для данного гена или участка ДНК).
При решении ряда научных и биотехнологических задач конструирование рекомбинантных ДНК требует также создания систем, в которых обеспечивается максимальная экспрессия клонируемого гена.
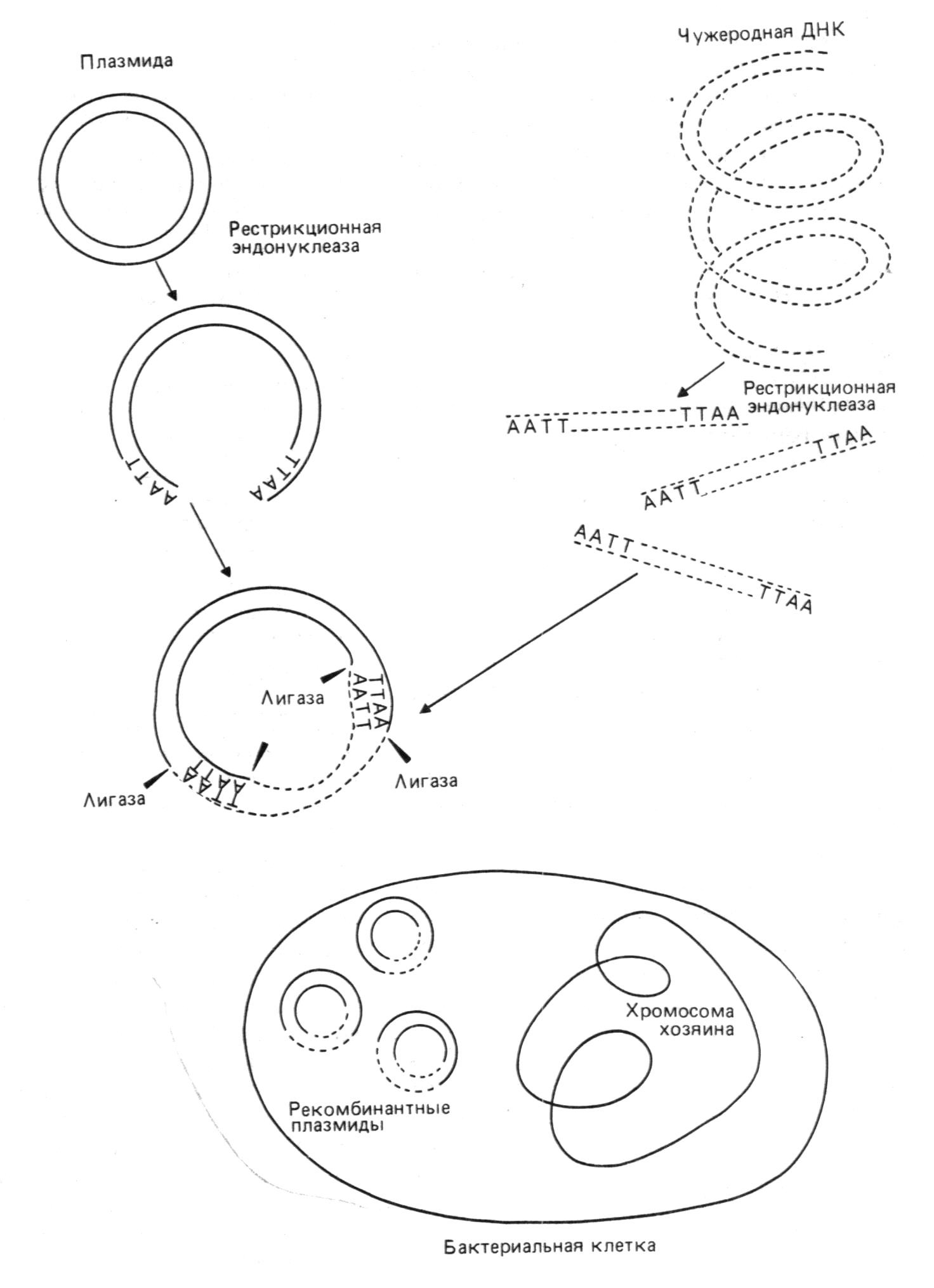
Существует три основных способа встраивания чужеродной ДНК в векторные молекулы. В первом случае 3'-концы фрагментов ДНК, среди которых находится интересующий нас участок ДНК (ген или его сегмент, регуляторный район), с помощью фермента терминальной нуклеотидилтрансферазы наращиваются гомополинуклеотидной последовательностью (например, поли (Т)). 3'-концы линейной формы векторной ДНК тем же способом наращиваются комплементарной ей гомополинуклеотидной последовательностью (то есть поли (А)). Это позволяет соединить две молекулы ДНК путем комплементарного спаривания искусственно полученных "липких" концов.
Во втором случае "липкие" концы создаются с помощью расщепления молекул ДНК (как векторной, так и содержащей интересующий нас фрагмент) одной из эндонуклеаз рестрикции (рестриктаз). Рестриктазы характеризуются исключительно высокой специфичностью. Они "узнают" в ДНК последовательность из нескольких нуклеотидных остатков и расщепляют в них строго определенные межнуклеотидные связи. Поэтому даже в ДНК больших размеров рестриктазы вносят ограниченное число разрывов.
Третий способ представляет собой комбинацию двух первых, когда липкие концы ДНК, образованные рестриктазой, удлиняются синтетическими последовательностями (рис. 1).
Концы фрагментов ДНК можно превратить в "липкие", наращивая их двутяжевыми олигонуклеотидами ("линкерами"), в состав которых входит участок узнавания рестрикта-
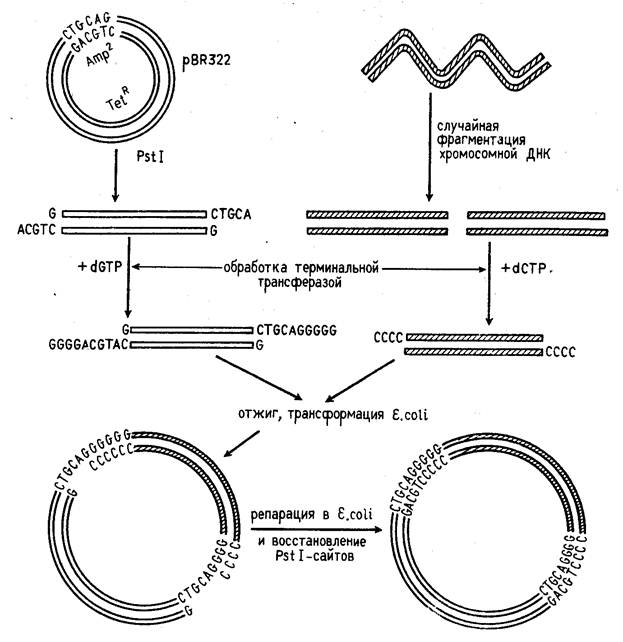
Рисунок 1. Схема конструирования рекомбинантной ДНК с помощью рестриктаз PstI и поли(G)- поли(С)-линкера.
зой. Обработка такого фрагмента данной рестриктазой делает его пригодным для встраивания в векторную молекулу ДНК, расщепленную той же рестриктаэой. Часто в качестве "линкера" применяются полинуклеотидные фрагменты, которые содержат специфические участки сразу для нескольких рестриктаз (их называют "полилинкерами").
После встраивания чужеродной ДНК в вектор их ковалентное сшивание осуществляется ДНК-лигазой. Если же размер бреши в рекомбинированной молекуле превышает одну фосфодиэфирную связь, она застраивается in vitro с помощью ДНК-полимеразы или in vivo с помощью репарирующих систем клетки.
1.2.2. ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ РНК
Получение рекомбинантных РНК обычно осуществляют методами ферментативного или химического лигирования РНК. Кроме того, недавно появилась принципиально новая возможность встраивания сегмента РНК в заданное положение других молекул РНК с помощью рибозимов.
Ковалентное сшивание отдельных сегментов РНК при получении рекомбинантных молекул, как правило, осуществляют с помощью Т4 РНК-лигазы. Т4 РНК-лигаза закодирована в геноме бактериофага Т4. Ее выделяют из клеток E.coli, зараженных этим фагом. Фермент сшивает друг с другом однотяжевые олиго- и полирибонуклеотиды. Для работы Т4 РНК-лигазы необходим источник энергии - аденозинтрифосфат. На рис. 2 приведена схема ферментативного лигирования двух коротких олигонуклеотидов. Как видно из этой схемы, акцептором в реакции лигирования служит полностью дефосфорилированный, а донором - полностью фосфорилированный по концевым нуклеотидным остаткам олигонуклеотид. Это предотвращает возможность сшивания однотипных олигонуклеотидов.
Эффективность ферментативного лигирования достаточно длинных полирибонуклеотидов сильно варьирует и ее трудно предсказать исходя только из нуклеотидной последовательности сегментов РНК. Наилучшие результаты получены в тех случаях, когда сшиваемые концы полирибонуклеотидов были пространственно сближены за счет комплементарного связывания соседних с ними участков РНК.
Недавно было установлено, что протяженные сегменты РНК (длиной в 200-300 остатков) могут быть с высоким выходом сшиты Т4 ДНК-лигазой. При этом "стыковка" сегментов осуществляется с помощью олигодезоксирибонуклеотида, комплементарного 3'-концу одного сегмента и 5'-концу другого.
Метод химического лигирования основан на активации концевой фосфатной группы одного из двух сшиваемых сегментов РНК водорастворимым карбодиимидом или
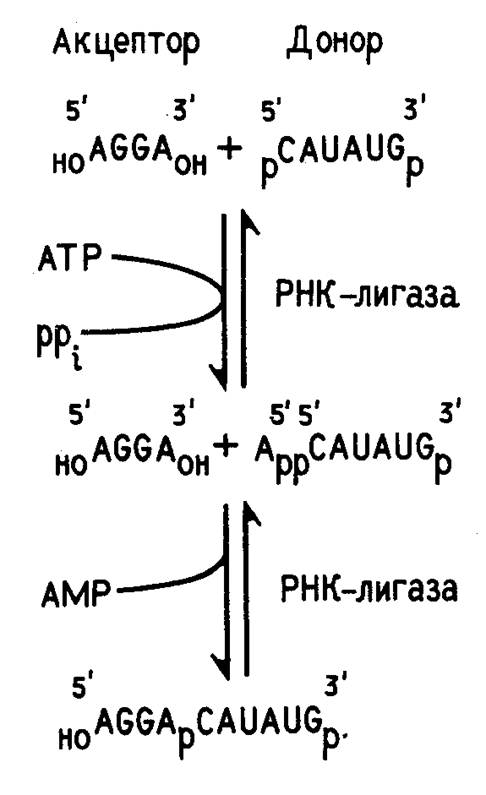
Рисунок 2. Схема сшивания двух олигорибонуклеотидов с помощью Т4 РНК-лигазы.
BrCN. В случае BrCN реакция протекает очень быстро и не сопровождается модификацией нуклеотидных остатков, хотя под действием карбодиимидов фосфодиэфирная связь образуется с более высоким выходом. Для того, чтобы обеспечить сближенность сшиваемых концевых нуклеотидных остатков в фрагментах РНК, было предложено использовать олигодезоксирибонуклеотиды, комплементарные обоим фрагментам в месте их стыка.
Химическое лигирование РНК, как правило, проходит с существенно меньшим выходом, чем ферментативное. Однако оно позволяет получать рекомбинантные РНК с необычными типами межнуклеотидной связи (например, пирофосфатной) и необычными нуклеотидными остатками в месте стыка двух фрагментов.
Получение рекомбинантных РНК с помощью рибозимов основано на обратимости реакции самосплайсинга (при отсутствии гуанозина или гуаниловых нуклеотидов). Это предоставляет возможность для встраивания интронной РНК в заданный участок другого сегмента РНК (рис. 3). Фрагмент РНК, в который производится встраивание, должен содержать нуклеотидную последовательность, идентичную нуклеотидной последовательности 3'-концевого участка 5'-экзонного района 26S РНК и соответственно комплементарную той нуклеотидной последовательности в интроне, которая отвечает за специфичность прямой реакции. Фрагмент, в который производится встраивание, берется в избытке.
В настоящее время описанная здесь цепь реакций может быть реализована только для интронной РНК, получаемой из предшественника 26S РНК тетрахимены. Однако можно думать, что конструирование новых рибозимов может существенно расширить возможности этого подхода.
Похожие работы
... олигонуклеотидов—одну полуавтоматическую, а вторую в комплексе с компьютером. В 1982 г. цена этих приборов на американском рынке составляла 36000—39500 долл.[2]. К открытиям связанным с достижениями генной инженерии нужно прибавить то, что огромный генетический «чертеж» многоклеточного существа просчитан полностью. Я думаю это можно назвать достижением века. После восьми лет работы многих ...
... генно-инженерных исследований. Многие из этих вопросов были подняты самими учеными активно работающих в данной области. В настоящее время большинство исследователей считали, что опасения касающиеся, генной инженерии, не имеют достаточно оснований, но многие этические проблемы остаются нерешенными и продолжают возникать новые. В прошлом генетика и медицинская генетика развивалась как относительно ...

... при постановке биологических или медицинских экспериментов. Составлять его должны ведущие специалисты в этой области и знающие предмет юристы. 3. Другие применения генной инженерии Несмотря на успехи ученых в генной инженерии в таких отраслях, как создание генно-модифицированных продуктов и клонировании животных и человека, они на этом не останавливаются. В целях сохранения военного ...
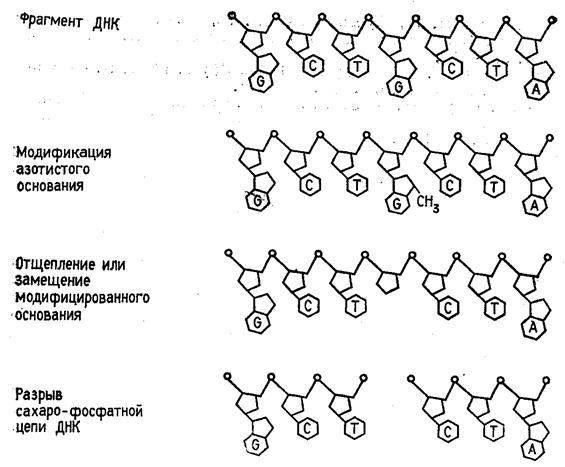
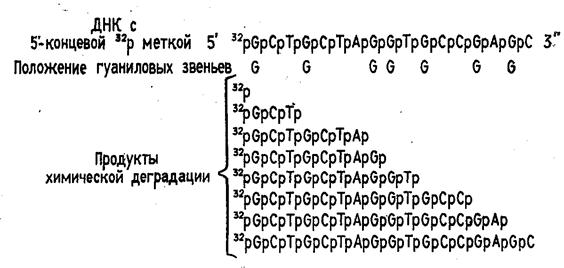
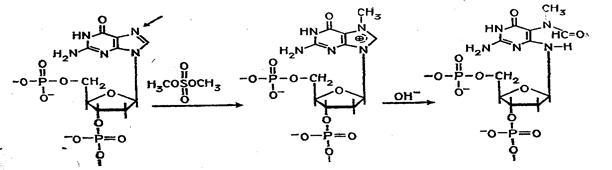
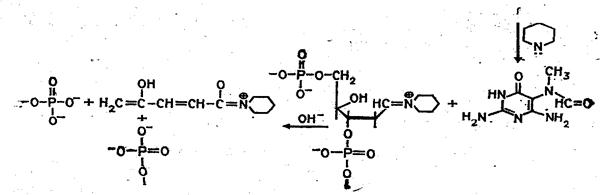
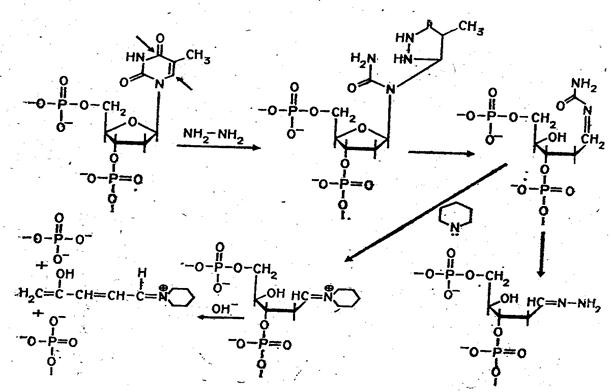
... (например, семейства генов) необходимо провести клонирование ПЦР-продуктов в векторах типа М13, в результате каждый фаг будет содержать только одну вставку. При прямом секвенировании смеси генов наблюдается несколько одинаково расположенных полос в разных дорожках геля. При амплификации же одного гена можно проводить прямое секвенирование, не прибегая к промежуточному субклонированию. Выбор ...
0 комментариев