Навигация
МОЛЕКУЛЯРНЫЕ ОСНОВЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ, РЕЦЕПТОРЫ
90264
знака
4
таблицы
11
изображений
6. МОЛЕКУЛЯРНЫЕ ОСНОВЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ, РЕЦЕПТОРЫ
Нейрон способен иметь до нескольких десятков тысяч межклеточных контактов, большинство из которых обеспечивается определенными морфологическими структурами — синапсами. Клеточную поверхность нейронов можно рассматривать как приемник разнообразнейших сигналов.
Чем выше степень эволюционной организации нервной системы, тем разнообразнее природа химических синапсов. Особенно это касается головного мозга высших млекопитающих, включая человека. Очевидно, химические синапсы оказались эволюционно более выгодными для передачи дискретных сигналов по сравнению с другими типами межклеточных контактов, поскольку на их 'основе возможна не только передача сигнала, ко и его разнообразная модуляция, в том числе гуморальными факторами. Основой восприятия нейроном химического сигнала в синапсе, а также ряда модулирующих влияний являются рецепторы.
Рецепторы представляют собой надмолекулярные образования, состоящие из белков, а также гликолипидных компонентов. Они способны под действием медиатора либо нашэсредственно изменять потоки ионов через мембрану, либо индуцировать образование вторичш>тх мессенджеров, которые, в свою очередь, меняют ряд свойста лейрона.
Межнейрональные химические синапсы подразделяются на два типа: возбуждающие и тормозные, причем.первые, как известно, способствуют генерации новых импульсов, а вторые приводят к снятию действия приходящих сигналов. Это деление определяется в значительной мере природой рецепторов. Известны случаи, когда один и тот же медиатор оказывает возбуждающее или тормозное действие в зависимости от природы рецептора.
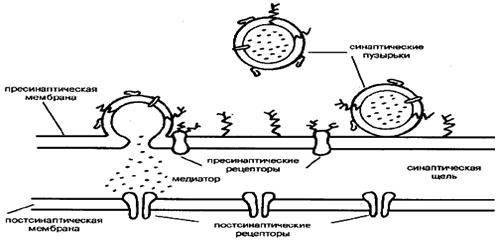
В зависимости от места положения синапсов их можно подразделить на сомато-аксональные, дентрито-аксональные, ден-трит-дентритные и др. Каждый из этих синапсов имеет свои особенности в функционировании. Схематически структура синапса может быть представлена следующим образом.
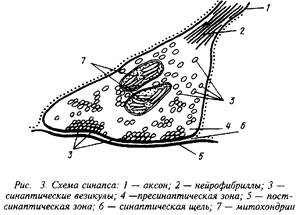
На рисунке хорошо видны утолщения, составляющие пре-синаптическую мембрану подходящего аксона, синаптическая щель и постсинаптическая мембрана. В пресинаптическом окончании находятся синаптические везикулы — хранилища запасов нейромедиатора в пресинаптическом нейроне. Постсинаптическая мембрана является носителем рецепторов. В ряде случаев сами рецепторы могут быть визуализированы при посредстве электронной микроскопии.
При распространении нервного импульса происходит деполяризация пресинаптической мембраны я изменение ионных токов. Наиболее важным событием в нервном окончании является мобилизация ионов Са, которые вызывают миграцию и открывание многочисленных синаптических везикул. Эти везикулы непосредственно связываются с участками пресинапса и открытие их приводит к высвобождению нейромедиатора и диффузии его в синаптическую щель. В терминали аксона сконцентрированы и ферменты синтеза медиатора, митохондрии для энергетического обеспечения этого процесса, системы белков-транспортеров, способствующих узнаванию и обратному захвату молекул нейромедиатора. Этот последний механизм, по-видимому, существенно экономит затраты на синтез готового нейромедиатора и участвует в регуляции срока его действия.
В отличие от биохимических процессов выброса нейроме-диаторов из пресинапса, имеющих общий характер, постсинап-тическое действие нейромедиатора прекращается самыми разнообразными способами: разрушением его определенными ферментами, либо быстрым поглощением из области синапса гли-альными клетками, либо обратным захватом его в пресинаптическую терминаль. В качестве примера существования разных механизмов утилизации нейромедиаторов из синапса можно привести механизмы процессов, происходящих в холинергиче-ском, глутаматергическом и ГАМК-ергическом синапсах.
Механизм разрушения ацетилхолина преимущественно связан с работой фермента — ацетилхолинэстеразы, который располагается на постсинаптической мембране и быстро гидроли-зует медиатор после взаимодействия с рецепторами. В глутаматергическом синапсе механизм удаления нейромедиатора заключается преимущественно в поглощении L-глутамата окружающими глиальными клетками. L-глутамат превращается в глутамин с помощью фермента глутаминазы, находящейся в глиальных клетках. В ГАМК-ергическом синапсе преобладает система обратного захвата медиатора.
7. ОБЩИЕ ВОПРОСЫ НЕЙРОРЕЦЕПТИИ
В биохимии и физиологии нервной системы длительное время доминировала точка зрения, согласно которой местом активных пластических изменений нейронов является пресинапти-ческая мембрана. Были представлены многочисленные свидетельства, касающиеся сдвигов количества квантов нейромедиатора в процессе обучения, памяти, выработки условных рефлексов и др. Постсинаптическим мембранам отводилась либо пассивная роль, либо они вообще не рассматривались в качестве активных участников событий в нервных клетках. Впоследствии стало ясно, что процессы, происходящие в плазматической или синаптической мембране нейрона, являются одними из ключевых для понимания интегративных функций ткани мозга, решения проблем обеспечения эффективного взаимодействия между нервными клетками. В последние десятилетия особое внимание было обращено на изучение структурных компонентов постсинаптических мембран, в частности нейрорецепторов. Исследования тонкой структуры и функции нейрорецепторов показали их важную роль в трансформации химического сигнала в биоэлектрические потенциалы и в передаче информации на внутриклеточные реакции, которые определяют метаболизм нервной ткани.
Следует отметить, что нейрорецепторы расположены как на мембранах нейронов, так и на мембранах глиальных клеток. Однако у последних они имеются в ограниченном наборе и числе. Рецепторные системы, расположенные на глиальных элементах, отличаются от нейрональных весьма важным моментом — они не способны продуцировать ответные реакции клеток в виде оперативных единиц информации. Иными словами, они не генерируют потенциалов действия. Как правило, глиальные клетки реализуют свое действие через внутриклеточные и межклеточные трофические регуляторные реакции, участвуя в метаболизме нейронглия.
Несмотря на огромное разнообразие клеточных рецепторов на мембране нейрона, их можно подразделять на два основных класса, которые различаются по механизмам действия и скорости проведения сигналов. Существуют быстродействующие ионотропные и медленнодействующие метаботропные рецепторы скорость действия первых составляет миллисекунды, в то время как у последних они находятся в секундно-минутном диапазоне. Время действия нейрорецепторов определяется структурной организацией рецепторных компонентов.
Быстродействующие рецепторы содержат в своей структуре ионный канал, открывающийся при контакте с нейромедиатором. Медленнодействующие рецепторы представляют собой комплекс из нескольких белков, которые при воздействии нейромедиатора последовательно меняют конформацию и в конечном счете активируют синтез или выход вторичного, уже внутриклеточного, медиатора. Эти два класса рецепторов обозначают нередко как рецепторы I и II класса. Для правильного восприятия терминологии целесообразно также указать, не рассматривая пока детали, что рецепторы класса II содержат в числе белков, передающих сигнал, так называемые G-белки. Их нередко упоминают, обозначая рецепторы этого класса.
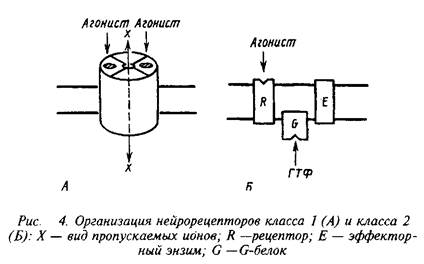
Кроме охарактеризованных выше двух классов рецепторов существуют еще три особые группы рецепторов, которые хотя и присутствуют в нервной системе, но пока представляются не связанными прямо со специфическими функциями последней. К ним относятся рецепторы, переносяшие свои лиганды через мембрану, рецепторы, обладающие собственной тирозинкиназной активностью и, наконец, своеобразная группа, которая при взаимодействии с лигандом претерпевает частичное протеолитическое расщепление. В настоящем руководстве мы не рассматриваем эти группы рецепторов.
К первому классу рецепторов принадлежат никотиновые рецепторы ацетилхолина, рецепторы ГАМКА, глицина, а также часть рецепторов глутамата и аспарагиновой кислоты. Рецепторы катехоламинов, серотонина, ГАМКВ и ряда пептидных соединений, а также мускариновые рецепторы ацетилхолина и некоторые из рецепторов глутамата относят ко второму классу. Последние типы рецепторов через систему вторичных посредников вызывают изменения в активности проте-инкиназ, способных фосфорилировать мембранные белки, включая ионные каналы.
Таблица 1
Структура, общий характер и функции рецепторов класса I
| Тип рецептора | Преимуществен -ная функция | Мг, кД | Субъединицы |
| Холинергический | Возбуждающая | 250 | а 40-50 Р 50-54 у 56-60 6 58-65 |
| ГАМКА | Тормозная | 230-260 | |
| Глутаматный | Возбуждающая | 240-400 | 90-10 |
| Глициновый | Тормозная | 246 | а 48 Р 58 |
Таблица 2
Масса белков рецепторов класса II
| Рецептор | Мг, кД |
| Ад рене р ги чески й | 58-80 |
| Глутаматер гически й | 90-110 |
| Холинергический | 85-105 |
| Дофаминовый | 72-94 |
| Опиатный | 53-65 |
| Серотониновьш | 67 |
Следует отметить, что в последние годы обнаружена группа нейрорецепторов, связь которых с ионными каналами осуществляется через G-белки, не сопряженные с перечисленными выше вторичными мессенджерами. Хотя в такую систему рецепции и не включены протеинкиназы, тем не менее участие G-белка в трансформации сигнала значительно увеличивает время действия по сравнению с нейрорецепторами класса 1.
Фундаментальным свойством всех нейрорецепторов является их лабильность и высокая скорость синтеза самого рецептора. Это свойство рецепторов конрастирует с более жесткой запрограммированностью синтеза белковых компонентов мембран, которая обычно наблюдается у других типов тканей. В нейронах развиты механизмы непрерывного синтеза рецепторов и их быстрой утилизации либо путем интернализации, либо с помощью пиноцитоза. Высокая скорость обновления нейрорецепторов обусловлена, по-видимому, необходимостью изменения "информационной емкости и "пропускной способности" нейрона. В этом случае генетический аппарат клетки способен, интегрируя всю приходящую информацию, "принять решение" путем перестройки синтеза белковых компонентов мембран. В этом скрыта одна из причин уникального свойства нейронов и нервной ткани в целом — пластичности.
Таким образом, основная роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов.
Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют альтернативу. С одной стороны, это изменение чувствительности к рецептору, с другой — увеличение или снижение количества активных рецепторов на мембране. Заслуживают внимания и гипотезы, касающиеся посттрансляционной модификации нейрорецепторов, которая позволяет изменить количественные параметры их функционирования.
Внимание к проблемам нейрорецептии со стороны биохимиков, фармакологов и физиологов обусловлено еще и тем, что причиной многих дисфункций нервной системы является нарушение целостности мембранных компонентов как нейронов, так и глиальных клеток. Отметим, что существующие успехи в лечении некоторых нервно-психических заболеваний связаны в большей мере с прогрессом в исследовании именно молекулярных свойств ряда рецепторов. Оказалось, что многие нейрорецепторы выполняют роль избирательных мишеней действия известных лекарственных препаратов. Исследования в этой области нейробиологии служат сейчас постоянным источником для целенаправленного поиска и создания новых классов фармакологических средств, обладающих улучшенными терапевтическими свойствами.
Похожие работы
... , накапливающие и хранящие медиатор вплоть до момента его освобождения в синаптическую щель. Мембрана синаптических пузырьков содержит разнообразные белки, предположительно участвующие в освобождении медиатора. 3. Материальная основа высшей нервной деятельности. Нервная система - совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и ...
... в состоянии продолжать жить, несмотря на указания совести о неправильности жизни…». На этом тезисе мы закончили предыдущую главу книги. Не развивая его, поскольку это не входит в достаточно узкую заявленную тематику оружия геноцида, обратимся к внутренним механизмам, которые включаются у людей после принятия психотропов. Мы знаем от чего люди убегают (от своей совести). Но за чем же люди гонятся? ...
... материалы хорошо описываются в рамках квантово-механической фононной Модели строения и функционирования клеточных мембран, что позволяет утверждать: “ФОНОН – КВАНТ биологической (клеточной) мембраны”. Модель пригодна для объяснения широкого круга наблюдаемых явлений. При этом наблюдаемые явления описываются в рамках единого понятийного аппарата и не требуют специфических допущений для описания ...
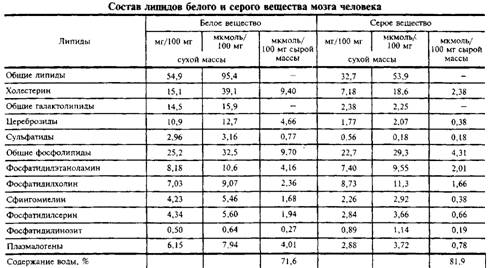
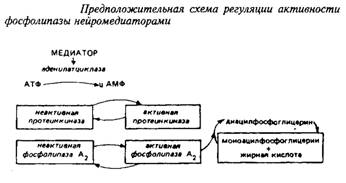
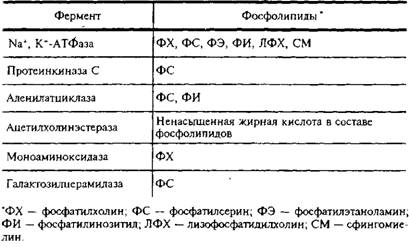
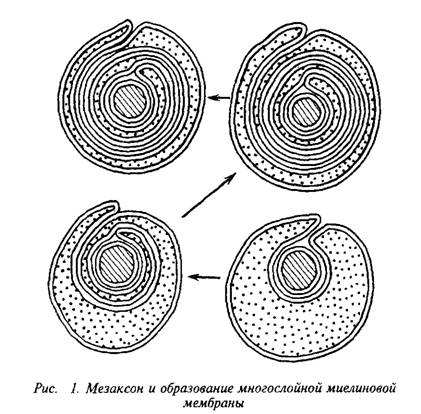
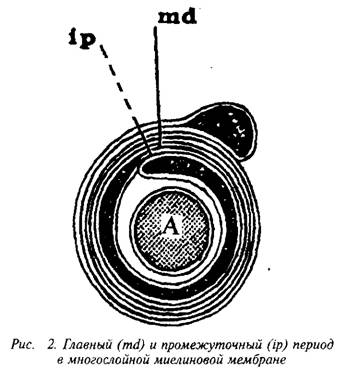
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
0 комментариев