Навигация
ОБЩИЕ ХАРАКТЕРИСТИКИ СТРУКТУРЫ И ФУНКЦИИ ИОНОТРОПНЫХ РЕЦЕПТОРОВ
90264
знака
4
таблицы
11
изображений
9. ОБЩИЕ ХАРАКТЕРИСТИКИ СТРУКТУРЫ И ФУНКЦИИ ИОНОТРОПНЫХ РЕЦЕПТОРОВ
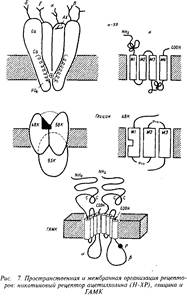
Основой всех ионотропных рецепторов является крупный белок, состоящий из пяти, реже четырех, белковых субъединиц. Молекулярные массы субъединиц варьируют обычно в пределах от 40 до 70 кД. Первичная структура белков различных ионотропных рецепторов обнаруживает высокую степень гомологии — от 20 до 60%, что указывает на общность эволюционного происхождения. Субъединицы рецептора пронизывают толщу клеточной мембраны, образуя ионный канал. Участки полипептидных полей субъединиц, выстоявшие над поверхностью клетки, служат для узнавания и взаимодействия с медиатором. Участки субъединиц, проходящие через толщу фосфолипидной мембраны и образующие собственно канал, характеризуются богатством гидрофобных неполярных аминокислотных остатков, обладающих высоким сродством к липидному окружению рецептора. Участки субъединиц, расположенные на внутренней поверхности мембраны, служат, во-первых, для взаимодействия с клеточными скелетными белками, ограничивающими их подвижность, и, во-вторых, являются мишенью для факторов, регулирующих активность рецептора в зависимости от ряда внутриклеточных процессов. Лучшим примером ионотропного рецептора служит рецептор ацетилхолина, представленный на рис. 7. Выстоящие над мембраной участки ионотропных рецепторов связаны нередко с углеводными компонентами.
Ионотропные рецепторы, например рецепторы гамма-ами-номасляной кислоты типа А, способны образовать большое количество подтипов за счет различного сочетания субъединиц. Существует более двух десятков подтипов ГАМКд-рецепторов благодаря различным комбинациям а-, р-, у- и5~субъединиц. В состоянии покоя каналы ионотропных рецепторов закрыты. При взаимодействии с медиатором происходит конформапионная перестройка субъединиц рецепторов и каналы открываются на несколько миллисекунду.
После активации рецепторные макромолекулы теряют на некоторое время чувствительность к медиатору. Наступает временная десенситизация.
Природа ионов, которые способен пропускать рецептор, определяется диаметром канала и характеристиками боковых радикалов аминокислотных остатков стенки канала. Никотиновые рецепторы ацетилхолина открывают дорогу ионам К* из клетки и ионам Na+ внутрь клетки; NMDA-глутаматные рецепторы наряду с одновалентными ионами, открывают путь внутрь клетки ионам Са+; ГАМКА и глициновые рецепторы пропускают внутрь клетки С1-ионы.
10. ОБЩАЯ ХАРАКТЕРИСТИКА МЕТАБОТРОПНЫХ МЕДЛЕННЫХ РЕЦЕПТОРОВ
Метаботропные рецепторы представляют собой сложную систему, состоящую, по крайней мере, из трех белков: 1) собственно рецепторного белка, связывающегося с нейромедиатором; 2) так называемого G-белка, модифицирующего и передающего сигнал с рецепторного белка и 3) белка-эффектора, который является ферментом, катализирующим образование внутриклеточного низкомолекулярного регулятора, так называемого вторичного мессенджера. Пример метаботропного рецептора представлен на рис. 8.
Собственно рецепторный белок — R-белок — представляет собой крупный полипептид, состоящий из 400-2 ООО аминокислотных остатков, N-конец пептида выстоит над поверхностью клетки, С-конец направлен внутрь клетки. Пептидная цепь семь раз пересекает клеточную мембрану, образуя соответственно по три петли над и под поверхностью мембраны. N-концевая последовательность нередко гликозилирована. Те части пептида, которые пронизывают фосфолипидную мембрану, состоят в значительной мере из неполярных аминокислот и как бы плавают в липидном слое. Медиатор, вступая во взаимодействие с внешними участками полипептида, меняет их конформацию и, в свою очередь, меняет положение плавающих внутри мембраны участков пептида. В конечном счете это ведет и к изменению конформации участков пептида, находящихся под мембраной. В этой конформации они приобретают способность контактировать со следующим белком комплекса — G-белком. Заметим, что активированный медиатором R-белок способен контактировать последовательно с многими десятками и сотнями молекул G-белка, переведя их, в свою очередь, в активное состояние. Иначе говоря, уже на этой стадии происходит усиление, амплификация сигнала.
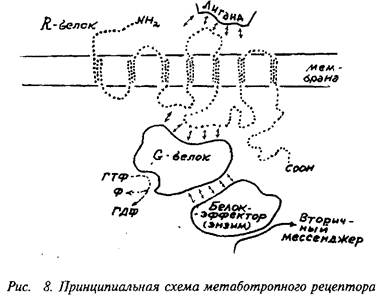
G-белок представляет собой олигомер, состоящий из 2-3 субъединиц с общей молекулярной массой порядка 60-100 кД, В неактивном состоянии G-белок обычно связан с молекулой ГДФ. При взаимодействии с активированным R-белком конфигурация G-белка меняется таким образом, что на место ГДФ становится ГТФ. Именно в состоянии комплекса с ГТФ G-белок способен быть активатором следующего компонента системы — фермента, образующего вторичный мессенджер. Активное состояние белка G-белка ограничено во времени тем, что связанные с ним ГТФ расщепляются до ГДФ, и G-белок при этом возвращается в исходное неактивное состояние. Расщепление ГТФ до ГДФ осуществляется самим G-белком, который является, как бы по совместительству, гуанозинтрифосфотазой.
Будучи в активном состоянии, G-белок активирует фермент. Этим ферментом может быть аденилатциклаза, катализирующая синтез цАМФ из АТФ, гуанилатциклаза, катализирующая синтез цГМФ из ГТФ, фосфолипаза С, отщепляющая фосфои-нозитол от фосфоинозитида мембраны. Активированный G-белок может выступать не только в роли фактора, усиливающего действие циклаз, но и в качестве их специфического ингибитора. Разновидности G-белка, выполняющие одну из этих функций, имеют обозначения: Gg-белок, стимулирующий активность циклаз, Gj-белок, подавляющий активность циклаз, и, наконец, О0-белок, активирующий фосфолипазу С. Это лишь главные и наиболее изученные разновидности G-белков. Вообще их число значительно больше. Отметим особо разновидности G-белков, которые, получив сигнал от метаботропного рецептора, передают его на тот или иной ионный канал. Строго говоря, это уже не метаботропный путь, а особая форма включения ионного канала. В отличие от ионотропных рецепторов здесь может, по-видимому, быть достигнута большая продолжительность действия и охват большего числа ионных каналов.
Так же как и стадии передачи сигнала с R-белка на G-белок, стадия активации фермента и стадия синтеза ферментом вторичных мессенджеров сопровождается дальнейшим усилением сигнала.
Похожие работы
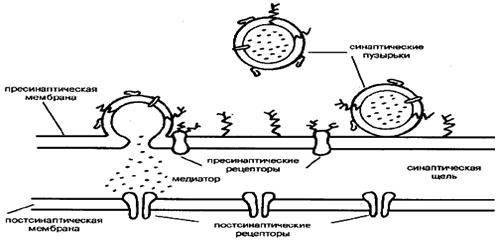
... , накапливающие и хранящие медиатор вплоть до момента его освобождения в синаптическую щель. Мембрана синаптических пузырьков содержит разнообразные белки, предположительно участвующие в освобождении медиатора. 3. Материальная основа высшей нервной деятельности. Нервная система - совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и ...
... в состоянии продолжать жить, несмотря на указания совести о неправильности жизни…». На этом тезисе мы закончили предыдущую главу книги. Не развивая его, поскольку это не входит в достаточно узкую заявленную тематику оружия геноцида, обратимся к внутренним механизмам, которые включаются у людей после принятия психотропов. Мы знаем от чего люди убегают (от своей совести). Но за чем же люди гонятся? ...
... материалы хорошо описываются в рамках квантово-механической фононной Модели строения и функционирования клеточных мембран, что позволяет утверждать: “ФОНОН – КВАНТ биологической (клеточной) мембраны”. Модель пригодна для объяснения широкого круга наблюдаемых явлений. При этом наблюдаемые явления описываются в рамках единого понятийного аппарата и не требуют специфических допущений для описания ...
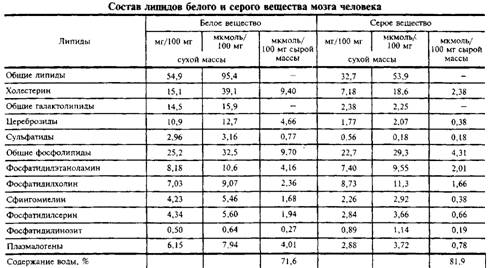
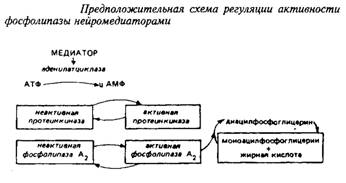
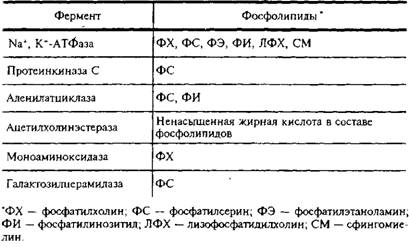
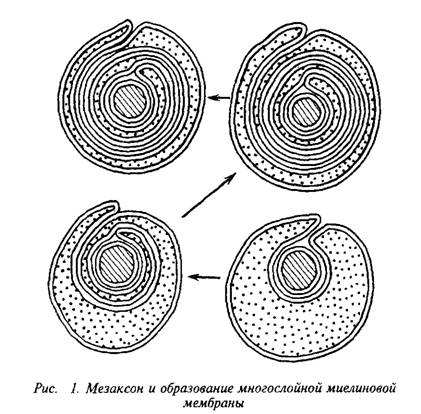
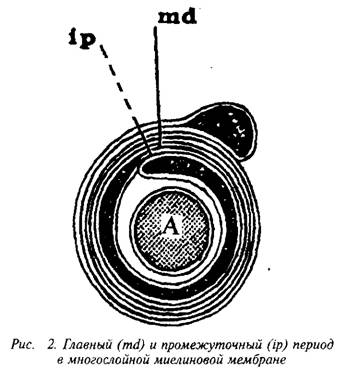
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
0 комментариев