Навигация
БИОХИМИЧЕСКИЕ МЕТОДЫ ИЗУЧЕНИЯ МЕХАНИЗМОВ НЕЙРОРЕЦЕПТИИ
90264
знака
4
таблицы
11
изображений
8. БИОХИМИЧЕСКИЕ МЕТОДЫ ИЗУЧЕНИЯ МЕХАНИЗМОВ НЕЙРОРЕЦЕПТИИ
Наиболее широко распространенным и разработанным методическим подходом для количественного анализа взаимодействия нейромедиаторов со своими рецепторами на мембране клетки является радиолигандный метод. Суть этого метода заключается в изучении параметров связывания радиоактивного лиганда с мембранно-связанными или изолированными рецепторными белками. В настоящее время существует хорошо развитая кинетическая теория рецептии и методы определения физико-химических параметров процесса образования комплекса лиганд-рецептор. Такой физико-химический анализ позволяет сделать определенные заключения о структуре активных центров нейрорецепторов, в частности, выяснить природу некоторых функциональных групп, которые ответственны за первую стадию взаимодействия лиганда с акцептором.
Для того чтобы кратко ознакомиться с количественной теорией взаимодействия веществ со своими рецепторами, рассмотрим простейшие условия, когда одна молекула лиганда взаимодействует с одним центром связывания:
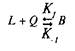
где L — лиганд; Q — центр связывания; В — комплекс лиганда со связывающим центром; К{ и K.j — кинетические константы. При динамическом равновесии скорость реакции образования комплекса В равна его скорости диссоциации, т.е. Vt = V_1? тогда концентрация вычисляется по формуле
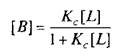
При этом предполагается, что L и Q взаимодействуют между собой по закону действующих масс, т.е. скорости реакций образования комплекса и его диссоциации прямо пропорциональны концентрациям компонентов в системе. Отношение констант прямой и обратной реакции называют константой сродства Кс. Она характеризует соотношение занятых и свободных участков связывания при данной концентрации лиганда. Обычно для описания параметров связывания используют величину, обратную константе сродства,—
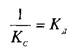
— константу диссоциации. Эта константа соответствует величине, при которой происходит насыщение 50% связывающих участков:
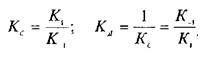
Если вместо константы сродства Кс использовать обратную ей величину Кд, то, подставив это значение в уравнение, характеризующее равновесную реакцию взаимодействия лиганда с рецептором, получим следующее уравнение:
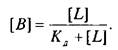
Приняв общее число рецепторов за 1, можно преобразовать уравнение к виду, аналогичному уравнению Михаэлиса, которое используется в энзимологии для описания кинетики обратимых ферментативных реакций:
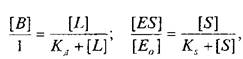
где — концентрация комплекса фермент-субстрат; —
концентрация субстрата и Ks — константа диссоциации комплекса; — исходная концентрация субстрата.
Согласно этим уравнениям зависимость величины эффекта от дозы лиганда или фермента описывается гиперболой. Чаще всего для работы пользуются графическим выражением зависимости эффекта не от концентрации, а от логарифма концентрации лиганда. Графически зависимость результатов может быть представлена разными способами, однако наиболее информативным способом расчета являются координаты Скэтчарда. Действительно, помимо равновесной константы связывания и общей концентрации центров связывания этот метод позволяет определить концентрацию свободного лиганда, соответствующую данной концентрации комплекса В. Константа диссоциации равна котангенсу угла наклона прямой. Отрезок на оси абсцисс от точки пересечения с прямой до начала координат соответствует максимальному уровню насыщения центров связывания.
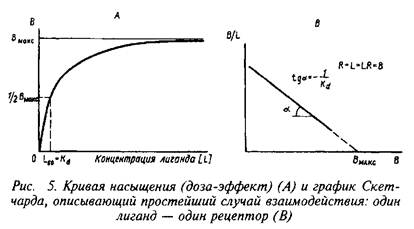
Таким образом, представление результатов равновесного связывания в координатах Скэтчарда дает информацию о характере протекающего процесса и позволяет определить важные параметры лиганд-рецепторного взаимодействия — константу диссоциации и концентрацию центров, способных образовывать комплексы с нейромедиатором.
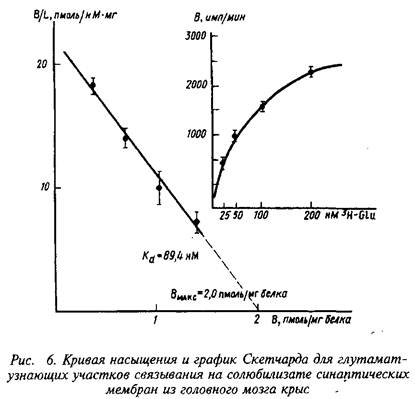
В качестве примера изучения рецепторного связывания нейромедиатора с белковыми компонентами на мембране нейрона приведем экспериментальные исследования глутаматных рецепторов радиолигандным методом. Так, исследования параметров связывания Н-глутамата с синаптическими мембранами, выделенными из коры больших полушарий головного мозга крыс, показали их зависимость от чистоты материала, способов хранения, условий проведения реакции связывания и др. При стандартизации всех указанных условий зависимость специфического связывания Н-глутамата с синаптическими мембранами имеет насыщающий характер. Представление экспериментальных данных в координатах Скэтчарда свидетельствует о наличии на мембранах однородной популяции участков связывания с Кд - 89,4 нМ и Вмакс = 2,0 пмоль/мг белка.
Значение количества центров связывания, выраженное в СРМ, пересчитывается в фмоль/мг белка по следующей формуле:
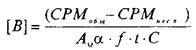 >
>
где А^ — молярная активность радиолиганда, Кю/моль; а — 2,210~ pacn/мин; f — эффективность счета; — разность счета связывания радиолиганда с рецептором в отсутствие и в присутствии немеченого радиолиганда; t — время счета; С — концентрация белка, мг.
Для того чтобы отличить эти параметры связывания от неспецифического связывания и поглощения глутамата другими участками мембраны, существуют дополнительные экспериментальные приемы, в том числе проведение реакции в присутствии разных катионов. Истинное рецепторное связывание глутамата является Na*-независимым процессом, в то время как поглощение и транспорт этого нейромедиатора другими участками синапса происходит в присутствии высоких концентраций ионов Na.
Далее возникает вопрос, соответствуют ли эти независимые участки связывания самого глутамата тем рецепторным компонентам на мембране нейрона, которые способны вызывать физиологический ответ клетки на данный медиатор. Оказалось, что сродство и константа диссоциации, полученные экспериментальным биохимическим методом, находятся в пределах физиологических концентраций действия L-глутамата на нейроны позвоночных. Такие показатели реакции связывания нейромедиатора, как насыщаемость и обратимость, соответствуют аналогичным свойствам глутаматного рецептора, регистрируемым с помощью электрофизиологических методов. Более того, чувствительность к ряду известных агонистов и антагонистов, таких как NMDA, каинат, квисквалат и другие, была сходна с физиологическими ответами. Следует упомянуть, что характер связывания нейромедиатора в присутствии ионов Na существенно отличается от рецепторного взаимодействия и коррелирует с параметрами высокоаффинного поглощения L-глутамата клетками, регистрируемыми физиологически. Все это иллюстрирует пути оценки параметров связывания нейромедиатора и специфические трудности, возникающие при такой оценке.
Одним из основных подходов к изучению молекулярных свойств нейрорецепторов является изолирование индивидуальных рецепторных белков, специфически связывающих нейромедиаторы или необратимо взаимодействующих с их антагонистами или бло-каторами. Так, прогресс в исследовании никотиновых холино-рецепторов был обусловлен обнаружением а-бунгаротоксииа, который оказался специфическим блокатором этого типа рецепторов и позволил выделить мембранные белки и очистить их на основе радиолигандного метода. Наличие таких приемов дает возможность разграничить хеморецепторные процессы от ферментативного и транспортного метаболизма нейромедиато-ров. Особенно это важно для изучения рецепторов аминокислотных медиаторов нервной ткани.
Изучение химической природы мембранных белков включает предварительное выделение, солюбилизацию, очистку и анализ очишенных компонентов. Причем применение классических методов структурного анализа для характеристики мембранных белков имеет свои сложности и особенности. Как правило, они обусловлены свойствами мембран и их компонентов, в частности, наличием липидных и гликолипид-ных структур. Проблемы, связанные с экстракцией белковых компонентов мембран, их очисткой и анализом, составляют специальный раздел мембранологии. Здесь будут рассмотрены лишь самые общие моменты.
Выбор метода солюбилизации зависит от цели исследования и имеет смысл только тогда, когда дает возможность сохранить нативные свойства рецепторного белка и исследовать его с помощью обычных биохимических подходов. Поэтому выбор со-любилизирующего агента на первом этапе может оказаться ключевым для анализа структуры и функции рецептора.
Существует целый ряд самых разнообразных солюбилизи-рующих агентов, пригодных для решения проблем мембранной биохимии. Наиболее надежными среди них являются неионные и ионные детергенты. В основе их действия лежит амфифиль-ная природа этих агентов, позволяющая им взаимодействовать и с гидрофильными, и с гидрофобными участками мембранных белков. Эффект детергента, разрушающего взаимосвязи в мембране, определяется двумя видами взаимодействия: детергент-белок и детергент—детергент. Большое значение имеет последнее взаимодействие, так как чем выше способность молекул детергента взаимодействовать друг с другом, тем меньше будет количество молекул, способных взаимодействовать с белками. Этот критерий мицеллообразования служит характеристикой детергента и его способности растворять те или иные белковые компоненты. Низкий коэффициент мицеллообразования характерен для мягких солюбилизирующих агентов, таких как тритон Х=Ю0, дезоксихолат натрия, дигитонин и другие, которые позволяют выделять нативные мембранные белки с сохранением их биологической активности. В то же время додецилсульфат натрия с высоким коэффициентом мицеллообразования обладает большой связывающей способностью и значительно повреждает нативную конформацию белков. Как правило, этот детергент используется при анализе субъединичной структуры макромолекул, так как легко разрушает межмолекулярные связи. Это свойство нередко применяется для определения молекулярной массы субъединиц белков при электрофорезе в присутствии ДСН.
Перед тем как приступить к дальнейшему выделению и изучению мембранных рецепторных белков, следует по возможности более полно удалить избыток детергента, поскольку он может оказывать нежелательное действие на биологическую активность и последующий физико-химический анализ структуры нейрорецептора.
Классические методы исследования мембранных белков, в том числе нейрорецепторов, включают практически все биохимические методы с учетом присутствия детергентов. Основным приемом специфического выделения ничтожно малых количеств нейрорецепторов является аффинная хроматография, которая позволила добиться впечатляющих успехов в изучении молекулярных свойств самых разнообразных типов нейрорецепторов.
Эффективность аффинной хроматографии зависит преимущественно от выбора лиганда или акцептора, который определяет природу выделяемого мембранного белка. Существенным фактором в этом случае является сродство лиганда к рецептору, и поэтому самыми эффективными лигандами оказываются специфические блокаторы или антагонисты нейрорецепторных белков. Иногда для выделения конкретного белка используют две или три ступени аффинной хроматографии на разных сорбентах и с разными лигандами. Получили широкое распространение методы иммуноаффинной хроматографии, в которых в качестве лиганда используется поликлональные или моноклональные антитела, полученные к компонентам рецептора.
Дальнейшее выделение и разделение фракций обычно осуществляют с помощью высокоэффективной жидкостной хроматографии, которая позволяет очищать индивидуальные компоненты мембранных белков. Причем обратнофазная хроматография дает уникальные возможности по разделению гидрофобных белков и пептидов. Нативность белковых компонентов рецепторов проверяют либо по лигандсвязьшающей функции, либо путем реконструкции их функции в разных модельных системах.
Одной из таких модельных систем, позволяющих контролировать ионтранспортные или ионселективные функции нейрорецепторов, служат липосомы. Способность липосом встраивать белки или целые рецепторные комплексы с сохранением их функциональной активности используется в мембранологии для моделирования функций белков "в чистом виде". В этом случае можно получать информацию о структурной организации компонентов, составляющих макромолекулу рецептора, и их внутренних перестройках в контролируемых условиях эксперимента.
В настоящее время разработано большое количество методов получения липосом, которые могут изменять фосфолипидный состав, заряд, "текучесть" или многослойность их компонентов. Размеры липосом могут варьировать от 25 нм до 100 мкм. Функцию белков, встраиваемых в липосомы, контролируют по динамике транспорта или накопления меченых ионов внутри липосом.
В последние годы исследователи возлагают особые надежды на иммунохимические способы идентификации структурных компонентов нейрорецепторов. Высокая специфичность антител и их способность узнавать разные антигенные детерминанты рецепторных комплексов широко используется для выяснения структурной организации нейрорецепторов и процессов их биосинтеза, включая генно-инженерные исследования. Иными словами, поли- и моноклональные антитела являются важным инструментом для изучения механизмов рецептии и общих вопросов нейробиологии.
Принципиальным решением множества проблем, связанных с применением антител, явилось создание новой гибридомной технологии, которая позволила получить моноклональные антитела. Эта техника была разработана в 1975 г. У.Келлером и А.Милстейном. Получаемые с помощью этого метода гибридные клетки синтезируют и выделяют в культуральную среду антитела, абсолютно одинаковые по своему сродству к той или иной антигенной детерминанте.
Гибридомная техника позволила получать самые разнообразные моноклональные антитела против химически индивидуальных антигенных детерминант на одной молекуле белка. В настоящее время моноклональные антитела широко используют для идентификации практически любых макромолекул, включая нейрорецепторы.
Следует подчеркнуть, что изучение структуры и функции нейрорецепторов и других мембранных компонентов нейрона является не самоцелью для нейробиологической науки. Важно понять, как молекулярные процессы, происходящие в нервных клетках, способны интегрировать самую разнообразную информацию и реализовать ее в виде сложных поведенческих, высших психических и эмоциональных реакций. Этот стратегический путь "от простого к сложному" получил в последние годы мощный импульс благодаря разработке принципиально новых способов прижизненной регистрации динамических биохимических реакций, происходящих в клетках головного мозга. Появились методы позитронно-эмиссионной томографии, ядерно-магнитного резонанса, гамма-сцинциграфии и другие, позволяющие прижизненно регистрировать системный метаболизм разных органов и тканей, включая головной мозг млекопитающих. Это создает предпосылки для успешного изучения нейрохимических основ формирования разнообразных функциональных состояний живого мозга человека, его наиболее сложной сферы деятельности —психической.
Одним из информативных методов является позитронно-эмиссионная томография, суть которой сводится к регистрации специальным устройством радионуклидных маркеров — меченых химических соединений, включающихся специфически в тот или иной метаболический процесс. Причем этот процесс может быть воспроизведен в виде томографических, т.е. объемных послойных изображений распределения позитронной метки по структурам и зонам головного мозга. Наиболее точная локализация достигается при использовании одновременно двух противостоящих детекторов, регистрирующих совпадающие лучи. В настоящее время уже существует хорошо разработанные приемы оценки функционального состояния головного мозга с помощью измерения локального метаболизма глюкозы, медиаторов, С02 и кислорода в процессе разнообразной деятельности индивида.
Важной особенностью метода, позволяющей его использовать в прижизненном исследовании деятельности мозга людей, является применение изотопов, излучения которых с учетом сроков распада безвредны для организма.
Сейчас получены четко различающиеся "карты" излучений при различных формах деятельности мозга человека, например восприятия слов, обдумывания слов, воспроизведения энграмм и др. Резкие различия регистрируются при воздействиях на мозг наркотиков и других психотропных агентов.
Естественно, что при дальнейшей разработке метода ПЭТ и его внедрении в клиническую нейробиологию возник вопрос о выборе адекватных маркеров, которые способны выявлять си-наптические реакции в нервной ткани. Ряд исследователей успешно работают с препаратами, которые позволяют визуализировать определенные нейрорецепторы и выявлять конкретные медиаторные пути, включающиеся в выполнение того или иного вида деятельности мозга человека. В клинике этот метод дает возможность проводить раннюю диагностику не только опухолевых новообразований, но и контролировать различные деструктивные процессы. Кроме того, применяя его, можно определять эффективность лечебного воздействия фармакологических средств и их правильный выбор для успешного лечения болезней мозга. В качестве иллюстрации к сказанному следует привести исследования, проводимые Вагнером и его коллегами по изучению вклада дофаминергических путей и их рецепторов в патогенез некоторых заболеваний. Выбор дофаминовых рецепторов был обусловлен их четкой локализацией в некоторых подкорковых экстрапирамидных структурах и известной их дисфункцией при двигательных расстройствах, паркинсонизме и шизофрении.
Предпосылкой для применения в ПЭТ агонистов и антагонистов дофаминовых рецепторов явились результаты радиоли-гандного связывания известных аналогов дофамина с синаптическими мембранами in vitro. Параметры связывания, константы диссоциации и количество связывающих участков сопоставляли с данными, полученными при ПЭТ, так как связывание радиофармпрепаратов с мембранами клеток, головного мозга in vivo имеет аналогичные закономерности. Расчет прижизненного взаимодействия нейрорецептор-лиганд имеет некоторые особенности, однако они учитываются непосредственно в программах компьютерного обеспечения. Наиболее удачными радио-лигандами для исследования дофаминовых рецепторов оказались антагонисты пС-метилспиперон и лУС-спироперидол. Поглощение и селективная избирательность накопления этих соединений в базальных ганглиях головного мозга коррелировали со степенью деструктивного процесса у больных паркинсонизмом. Эти исследования подтвердили гипотезу о первичной вовлеченности нигро-стриарных структур в регуляцию двигательных функций.
Дальнейшее развитие регистрирующих устройств ПЭТ, увеличение степени разрешения приборов на микроуровне и поиск новых избирательных радиофармпрепаратов могут открыть уникальные перспективы прижизненного изучения динамических биохимических процессов, происходящих в ткани мозга при выполнении сложных видов деятельности.
Похожие работы
... , накапливающие и хранящие медиатор вплоть до момента его освобождения в синаптическую щель. Мембрана синаптических пузырьков содержит разнообразные белки, предположительно участвующие в освобождении медиатора. 3. Материальная основа высшей нервной деятельности. Нервная система - совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и ...
... в состоянии продолжать жить, несмотря на указания совести о неправильности жизни…». На этом тезисе мы закончили предыдущую главу книги. Не развивая его, поскольку это не входит в достаточно узкую заявленную тематику оружия геноцида, обратимся к внутренним механизмам, которые включаются у людей после принятия психотропов. Мы знаем от чего люди убегают (от своей совести). Но за чем же люди гонятся? ...
... материалы хорошо описываются в рамках квантово-механической фононной Модели строения и функционирования клеточных мембран, что позволяет утверждать: “ФОНОН – КВАНТ биологической (клеточной) мембраны”. Модель пригодна для объяснения широкого круга наблюдаемых явлений. При этом наблюдаемые явления описываются в рамках единого понятийного аппарата и не требуют специфических допущений для описания ...
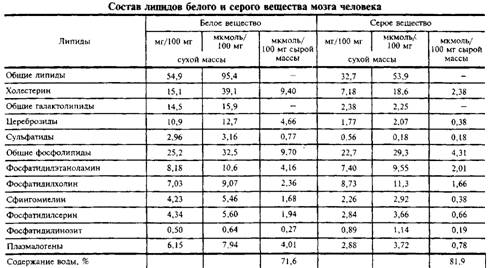
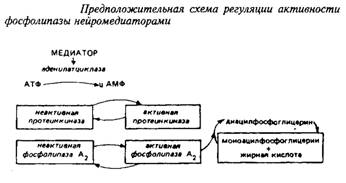
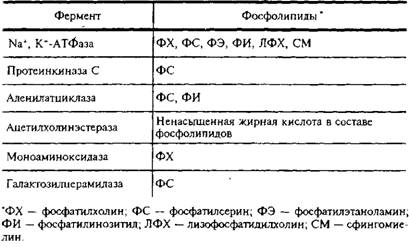
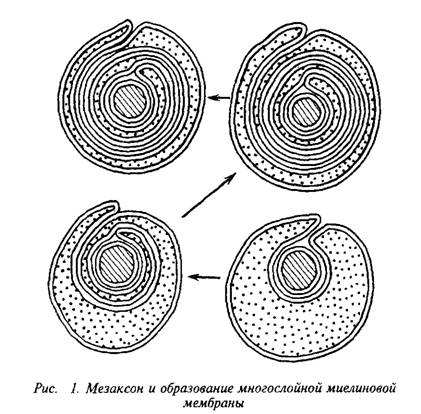
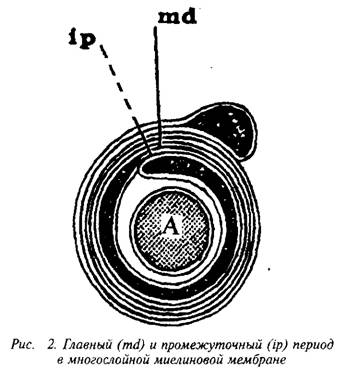
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
0 комментариев