Навигация
Молекулярные основы эволюции, дифференцировки развития и старения
28505
знаков
0
таблиц
1
изображение
Молекулярные основы эволюции, дифференцировки развития и старения
Известно, что некоторые фрагменты ДНК могут перемещаться с одного места на другое в пределах одной хромосомы или встраиваться в другую хромосому.
Существование прыгающих генов впервые было показано Б. Мак-Клинток при изучении генетики кукурузы. Она выяснила, что элементы регуляторного гена перемещаются в геноме с одного места на другое и влияют на экспрессию генов, приводя к появлению фенотипических вариаций. Эта работа не привлекала внимания в течение 30 лет, пока исследования на E. Coli не подтвердили способность генов к передвижению и внедрению в другое место в геноме, а Б. Мак-Клинток была присуждена Нобелевская премия.
Способные к перемещению последовательности ДНК получили название транспозоны (Tn‑элементы) или прыгающие гены.
Изменение положения какого-то сегмента ДНК относительно окружающих его последовательностей происходят повсеместно в разных вариантах, однако большинство из них наблюдается относительно редко. Многие геномные перестройки происходят в результате гомологичной рекомбинации между аллельными последовательностями и не затрагивают соседних сегментов ДНК. Вообще же говоря, структура генома вполне стабильна. Это и не может быть иначе, поскольку необходимо поддерживать жизнеспособность особей и видов.
Многие геномные перестройки не запрограммированы, они не связаны с каким-то специфическим влиянием на экспрессию генов и в них есть элемент случайности. Случайными могут быть частота таких событий, сами сегменты ДНК или то и другое. Примерами таких довольно редких событий служит транспозиция последовательностей ДНК из одного геномного локуса в другой или дупликация и последующая амплификация сегментов ДНК. Однако сходные транспозиции и амплификации могут быть сопряжены также с неслучайными, запрограммированными изменениями. Такие запрограммированные события играют ключевую роль в регуляции экспрессии некоторых генов во время дифференцировки и развития определенных типов клеток.
Типы мобильных элементов
Различают несколько типов мобильных ДНК-элементов, но все они обладают следующими общими свойствами:
- несут ген (или несколько генов), необходимый для транспозиции (ген транспозазы – фермента участвующего в перемещении);
- во-вторых, на концах содержат специфические взаимно инвертированные повторяющиеся последовательности, также необходимые для транспозиции
- сами транспозирующиеся элементы не кодируют никаких существенных для организма функций, однако часто содержат специфические гены, например ген устойчивости к антибиотикам. Транспозиция этих элементов, как правило, сопровождается сильными мутагенными эффектами.
- К транспозирующимся элементам относят те, транспозиция которых протекает без участия обратной транскрипции
Транспозирующиеся элементы прокариот
1. Инсерционные последовательности (IS – insertion sequences – последовательности вставки).
- Это сегменты‚ способные как целое перемещаться из одного участка локализации в другой (рис 73 Коничев).
- Ип содержат лишь те гены‚ которые необходимы для их собственного перемещения – транспозиции.
- на обоих концах каждого из них всегда имеются инвертированные повторы (рис. 10.4), которые также необходимы для транспозиции. Нуклеотидные последовательности этих повторов различны для разных IS и варьируют по длине от 10 до 40 п. н.
- Их перемещения в новые геномные локусы часто приводят к мутациям, заключающимся в прерывании регуляторных и кодирующих участков‚ а промоторные элементы внутри самих IS могут влиять на экспрессию близлежащих генов.
- Частота транспозиций у разных элементов неодинакова и составляет 10–5–10–7 на поколение.
- При транспозиции IS в новое положение исходный IS – элемент остается на прежнем месте; т.е., инсерция сопровождается точным синтезом второй копии и зависит от репликативных функций хозяина.
2. Транспозоны.
– Т. называют сегменты ДНК‚ обладающие теми же свойствами‚ что и ИП‚ но содержащие также гены‚ не имеющие отношение непосредственно к транспозиции
а) сложные транспозоны
– Часто IS – элементы длиной примерно 1 т. п. н. входят в состав более сложных транспозонов (прозрачка 1).
– Их длина может достигать тысяч пар оснований, а центральные области могут содержать разнообразные гены, (например‚ гены устойчивости к антибиотикам‚ гены токсинов или гены дополнительных ферментов клеточного метаболизма)
– С обеих сторон эта центральная область фланкируется одинаковыми IS‑элементами (от IS 1 до IS 10) – IS – L и IS – R с левой и правой сторон соответственно.
– Вся информация, необходимая для перемещения сложного транспозона, содержится в его IS – части; это та самая информация, которая используется IS‑элементом как таковым.
б) Простые транспозоны.
– это транспозоны не связаны ни с какими IS – элементами:
- информация, необходимая для транспозиции, закодирована в них самих.
- В качестве примера можно привести транспозон ТnЗ (прозрачка 3), который содержит инвертированные концевые повторы длиной 38 п. н., I R-L и I R-R, необходимые для транспозиции.
- Между этими повторами находятся три гена: два из них кодируют транспозиционные белки, третий – β-лактамазу (ген атр, не связан с транспозицией).
- Кроме того, имеется некодирующий участок длиной 170 п.н., содержащий промоторы генов транспозиционных белков, а также специфический участок, называемый res и тоже необходимый для транспозиции.
Сайты-мишени
Это сегменты ДНК, в которые встраиваются Мэ
– Одни транспозирующиеся элементы довольно разборчивы и охотнее встраиваются в короткие геномные сегменты, гомологичные концам самого мобильного элемента.
– Другие менее капризны и не отдают явного предпочтения никаким сайтам-мишеням, хотя наблюдается некоторая тенденция к встраиванию их в АТ-богатые участки.
– Независимо от типа Мэ. его встраивание в новый генетический локус обычно сопровождается дупликацией короткого участка ДНК в в сайте-мишени.
– Эти дуплицированные сегменты затем фланкируют встроившийся элемент.
– Почти обязательная дупликация сайтов-мишеней указывает на то, что при различиях в механизме большинство вставок происходит с образованием в потенциальных сайтах-мишенях смещенных одноцепочечных разрывов, как это показано на рис. 10.1. Сайт-мишень в геноме, содержащий мобильный элемент, условно называют «заполненным», а не содержащий такового – «свободным».
Типы транспозиций:
Для простоты рассмотрим транспозицию из одного генома в другой (например, из плазмиды в бактериальный геном или обратно). Внутримолекулярная транспозиция протекает более сложно.
Похожие работы
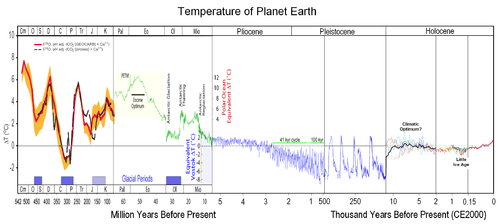
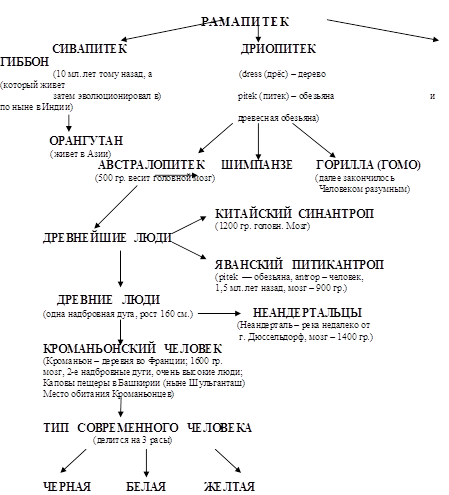

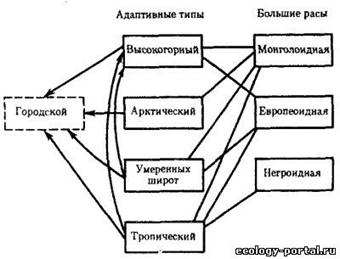

... важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии. Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную (" ...
... в области геронтологии. Это общая теория старения. Само собой разумеется, речь идет только об эскизах. Собственно это еще не общая теория старения, а только её очерк. Благодаря тому, что и как я сделал, моя скромная работа весьма отличается от известных до сих пор трудов по геронтологии. К сожалению, я не могу сразу предложить точные дефиниции, но, по крайней мере, имею возможность сделать ...
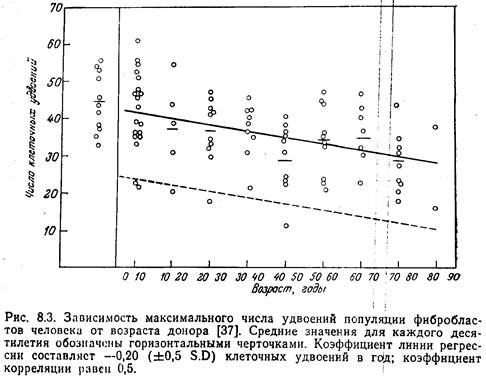
... - ядре, митохондриях, мембранах и др; в разных типах клеток- нервных, секреторных, иммунных, печёночных и др. Темп возрастных изменений определяется соотношением процессов старения и витаукта. Механизмы могут быть разделены на 2 группы. 1. Генотипические- генетически запрограммированные механизмы: а) система антиоксидантов, связывающая свободные радикалы; б) система микросомального ...

... , а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны. 3. Центральная догма и принцип Вейсмана принимаются. 4. Эволюция осуществляется путем изменения частот генов. 5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и ...
0 комментариев