Навигация
3. Ретропозоны
В геноме млекопитающих, птиц, амфибий и насекомых обнаруживаются ретропозоны – внедрившиеся в геном ДНК-копии, синтезированные на разных типах клеточных РНК, как на матрицах. Структура ретропозонов позволяет с уверенностью говорить об участии обратной транскрипции в процессе их образования. Таким образом, наряду с переносом информации от ДНК к РНК осуществляется и обратный процесс – возвращение ее в геном в виде ретропозонов. У млекопитающих ретропозоны составляют более 10% ДНК, следовательно, мощность встречного потока информации от РНК к ДНК может быть существенной, по крайней мере при оценке его во временном эволюционном масштабе.
Различают разные типы ретропозонов, среди них:
А) Псевдогены. Это копии генов, лишенные интронов и обладающие последовательностью поли-дА-поли-дТ на 3'-фланге. Матрицей при их копировании послужила «процессированная» полиаденилированная мРНК. Рамки трансляции таких «генов» часто «испорчены» стоп-кодонами и делециями. Они, как и проретровирусы, ограничены короткими прямыми повторами, представляющими собой дупликацию геномной последовательности в области сайта внедрения ДНК-копии, образованной на мРНК.
Б) Ретрогены. Если ретропозон правильно процессирован, ДНК-копия содержит открытую рамку считывания и возможна его экспрессия, то такие ретропозоны называют ретрогенами.
Большую массу повторяющихся элементов позвоночных представляют ДНК-копии клеточных РНК, кодирующих белки неизвестной природы, а также аномально процессированных клеточных транскриптов тРНК, 7S РНК и UPНК. Они представляют собой длинные и короткие повторы. Семейства повторяющихся последовательностей часто получают свои названия по расщепляющим их рестриктазам (например, короткие Alu‑повторы или длинные Крn‑повторы в геноме человека).
Каждый вид млекопитающих обладает собственным главным семейством длинных повторов, отличающих его от других видов. Предполагают, что разные семейства длинных повторов возникли сравнительно недавно, уже после эволюционной дивергенции видов млекопитающих.
Короткие повторы, например повторы Alu – семейства у человека, составляют 3 • 105, что соответствует 5 – 6% массы ДНК клетки. Их нуклеотидная последовательность гомологична последовательности отдельных участков 7S РНК. Такие повторы рассеяны по геному, их обнаруживают в интронах, на 5'-флангах генов, в составе З'-нетранслируемых участков гена. Между особями одного вида иногда наблюдаются различия (полиморфизм) в структуре генов и их ближайшего окружения, обусловленные внедрением коротких или длинных повторов. В геноме существует множество неактивных дефектных копий этих элементов, они не способны кодировать транспозазу или обратную транскриптазу. С другой стороны, они сохраняют способность к перемещениям, если в случае транспозонов имеются инвертированные повторы, узнаваемые транспозазой, а в случае ретротранспозонов сохранены промотор и возможность транскрипции элемента. Множество таких дефектных копий начнут перемещаться, если ферменты, ответственные за перемещение, будут кодироваться другими полноценными элементами. В геноме человека источником активной обратной транскриптазы является ретротранспозон L1, число копий которого достигает 100 тыс. Из них активно перемещаются 30 – 60 тыс., остальные, поврежденные, перемещаться не могут.
Мобильные генетические элементы могут принимать участие в перестройках хромосом. Наличие в хромосомах нескольких одинаковых по нуклеотидной последовательности копий подвижного элемента позволяет в редких случаях осуществить рекомбинацию по районам их локализации. В результате неравного кроссинговера возникают делеции отдельных участков или, наоборот, дупликации (рис. 98).
Неравный кроссинговер по районам локализации транспозона mariner (моряк) у человека приводит к делеции участка в коротком плече 17‑й хромосомы. Если это событие происходит в зародышевой клетке при созревании гамет, то хромосома с делецией будет передана потомкам. Это приводит к наследственным заболеваниям нервной системы – невропатиям и параличам. Внутрихромосомная рекомбинация между двумя элементами приводит к инверсии – повороту участка хромосомы на 180°. Инверсия может быть вредна для организма, а может способствовать эволюции генома, поскольку помогает передать потомству случайно сложившееся благоприятное сочетание генов, препятствуя кроссинговеру.
Ретротранспозоны с ДКП участвуют в ликвидации двунитевых разрывов ДНК. Обычно такой разрыв залечивается с помощью гомологичной молекулы ДНК, например сестринской, только что регошцированной нити. Участие реплицирующейся ДНК ретро-транспозонов с ДКП в процессе заживления двунитевых разрывов обнаружено у дрожжей (рис. 99). Брешь в двунитевой спирали, заделанная ретротранспозоном, сохраняет целостность хромосомы, но изменяет ее нуклеотидную последовательность. Если район хромосомы, где была брешь, не содержит существенного гена, то клетка остается жизнеспособной.
Подвижные элементы не стоит рассматривать только как «эгоистическую» ДНК, паразитирующую на ДНК клетки-хозяина. Они приспособлены и для нужд генома клетки: способствуют сохранению структурной целостности хромосомной ДНК; внедряясь в гены, не только инактивируют, вызывая мутации, но и меняют характер их экспрессии. В связи с этим подвижные генетические элементы, будучи важными факторами изменчивости генов и участвуя в перестройках структуры хромосом, имеют огромное значение в процессах эволюции геномов.
Похожие работы
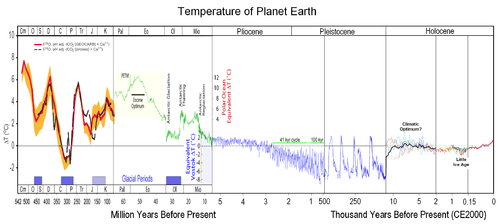
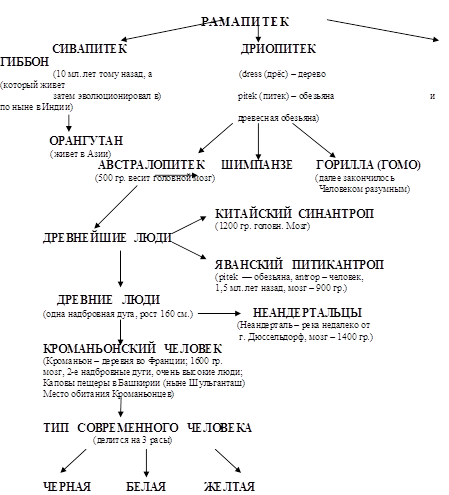

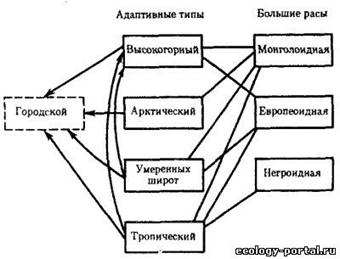

... важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии. Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную (" ...
... в области геронтологии. Это общая теория старения. Само собой разумеется, речь идет только об эскизах. Собственно это еще не общая теория старения, а только её очерк. Благодаря тому, что и как я сделал, моя скромная работа весьма отличается от известных до сих пор трудов по геронтологии. К сожалению, я не могу сразу предложить точные дефиниции, но, по крайней мере, имею возможность сделать ...
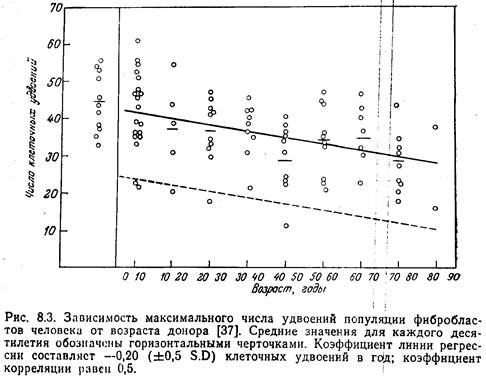
... - ядре, митохондриях, мембранах и др; в разных типах клеток- нервных, секреторных, иммунных, печёночных и др. Темп возрастных изменений определяется соотношением процессов старения и витаукта. Механизмы могут быть разделены на 2 группы. 1. Генотипические- генетически запрограммированные механизмы: а) система антиоксидантов, связывающая свободные радикалы; б) система микросомального ...

... , а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны. 3. Центральная догма и принцип Вейсмана принимаются. 4. Эволюция осуществляется путем изменения частот генов. 5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и ...
0 комментариев