Навигация
Коинтеграционная или репликативная транспозиция (поскольку происходит полная дупликация элемента.)
28505
знаков
0
таблиц
1
изображение
1. Коинтеграционная или репликативная транспозиция (поскольку происходит полная дупликация элемента.)
- донорный геном, который несет транспозирующийся элемент, сливается с реципиентной молекулой ДНК (прозрачка 4).
- Образовавшийся коинтеграт содержит всю донорную и реципиентную ДНК, а также по одной копии транспозирующегося элемента в местах сочленения этих ДНК.
- Коинтеграция включает разрыв исходных фосфодиэфирных связей и образование новых, дупликацию всего элемента и дупликацию сайта-мишени.
- Коинтеграт может затем разрешаться с образованием двух исходных ДНК, каждая из которых несет копию транспозированного элемента. Для разделения коинтеграа необходимодействие продукта гена tnp R‚ называемого резолвазой (от англ. Resolution – разрешение)‚ которая разрезает коинтеграт на исходные репликаторы.
- При транспозиции путем коинтеграции используется не только информация, закодированная в самом элементе, но и репликативные функции клетки.
2. Простое встраивание или консервативное (или нерепликативным, поскольку дупликации как таковой не происходит)
- Транспозирующийся элемент перемещается в новый геномный локус, при этом никаких других перестроек, кроме дупликации сайта-мишени, не происходит (прозрачка 4).
- Некоторые транспозирующиеся элементы, например ДНК фага Мu, участвуют как в коинтеграции, так и в простом встраивании.
3. Существует еще одна весьма привлекательная модель, согласно которой:
- на основе общей промежуточной структуры осуществляется транспозиция любого из двух типов (рис. 10.9).
- В соответствии с этой моделью может происходить транспозиция ДНК фага Мu и других мобильных элементов.
- На рисунке, иллюстрирующем модель, и ДНК донора, и ДНК реципиента представлены в кольцевой форме, поскольку в экспериментах in vitro, поставленных для проверки справедливости данной модели, использовалась кольцевая плазмидная ДНК;
- мобильным элементом в донорной ДНК служила модифицированная ДНК фага Мu. In vivo доноры и реципиенты, например плазмиды и хромосомы Е.соli, также находятся в кольцевой форме.
- Согласно данной модели, транспозиция начинается с образования одноцепочечных разрывов в кольцевых донорной и реципиентной ДНК.
- У донора такие разрывы происходят с обоих З'-концов элемента, у реципиента – в сайте-мишени с образованием 5'-выступов.
- Затем разрезанные концы мобильного элемента соединяются с концами реципиента, и образуется общий промежуточный продукт.
- Если в цепях, противоположных по отношению к разрезанным первоначально, никаких дополнительных разрывов не происходит, то имеет место простое встраивание.
- В результате заполнения бреши и лигирования происходит дупликация сайта-мишени.
- Таким способом может осуществляться нерепликативная транспозиция Тn 10.
- Эту реакцию катализирует транспозаза Тn 10, которая ускоряет разрезание на концах Тn 10, а также, по-видимому, воссоединение разорванных концов.
- При репликативной транспозиции общий промежуточный продукт претерпевает совсем другие превращения.
- интермедиат по своей структуре аналогичен кольцевой ДНК с двумя репликативными вилками.
- Если репликация начинается в каждой вилке, то конечный продукт представляет собой коинтеграт-кольцо, содержащее и донорную, и реципиентную ДНК, а также копии транспозирующегося элемента, реплицированные полуконсервативным путем.
- При разрешении коинтеграта в конце концов осуществляется транспозиция новой копии мобильного элемента в другой сайт-мишень. Разрешение может происходить путем гомологичной рекомбинации между двумя копиями транспозирующегося элемента с участием рекомбинационного аппарата Е.соli. (прозрачка 6).
Генетическая изменчивость бактерий при транспозиции
Мобильные элементы вызывают генетическую нестабильность поблизости от участка своей локализации, особенно в процессе репликативного механизма транспозиций.
В зависимости от того, как внесены разрывы в ДНК-мишень, получится либо делеция (Выпадение участка хромосомы из ее внутренней области), либо инверсия (перестановка) генетического материала между местом расположения транспозона и мишенью его перемещения. В связи с этим интересно отметить, что хромосомы родственных видов бактерий отличаются друг от друга многочисленными перестройками именно этого типа. По-видимому, мобильные элементы сыграли существенную роль в дивергенции и видообразовании бактерий.
1) Встраивание IS‑элементов поблизости от молчащего гена может приводить к его активации за счет транскрипции с промотора IS – элемента, т.е. изменяется регуляция бактериального гена (пр. 8а).
2) Очень важно, что мобильные элементы служат подвижными участками гомологии, гомологическая рекомбинация между которыми может приводить к дупликациям генов (рис. 76 Коничев). Считается, что дупликация – один из основных путей эволюционного возникновения новых функций. Действительно, «лишняя» копия гена выходит из-под давления естественного отбора и получает возможность накапливать изменения. Чаще всего это приведет к утрате какой бы то ни было функции, но иногда может получиться ген с новыми функциями.
3) Нельзя забывать и тот факт, что клетка может получить селективное преимущество за счет приобретения в составе транспозона гена, который сам по себе способен оказаться выгодным для бактерии в определенных условиях. Действительно, на транспозонах «путешествуют» гены устойчивости к различным бактериальным ядам, в том числе к тяжелым металлам и антибиотикам, гены дополнительных метаболических путей, позволяющие использовать необычный источник углерода, наконец, гены некоторых токсинов, делающие бактерии патогенными и позволяющие им тем самым существенно изменить образ жизни. Сказанное в равной степени относится и к плазмидам, поскольку большинство полезных для клетки-хозяина плазмидных генов находится в составе транспозонов.
4) Если элемент встроен в один из генов полицистронного оперона, то встроенный элемент может влиять на экспрессию последующих генов, либо останавливая транскрипцию в сайте терминации, находящемся внутри его самого (прозрачка 8,6), либо подавляя трансляцию дистальных кодирующих последовательностей мРНК (прозрачка 8, в)
5) Когда один и тот же геном служит и донором, и реципиентом при образовании коинтеграта (внутримолекулярная транспозиция), способность мобильных элементов вызывать перестройки в соседних участках ДНК приводит к другим мутационным эффектам. Все происходящие при этом события, включая делеции и инверсии, связаны с соединением одного или двух концов мобильного элемента с новыми последовательностями ДНК (рис. 10.13); детали этих процессов до конца не установлены.
Подвижные генетические элементы у эукариот
Существенную часть генома эукариот (10–30%) составляют повторяющиеся последовательности, имеющие определенную структурную организацию и способные перемещаться в геноме как в пределах одной хромосомы, так и между хромосомами. Они получили название подвижных генетических элементов. Элементы включают в свой состав от тысячи до десятков тысяч нуклеотидных пар.
Наибольшее количество подвижных элементов обнаружено в геноме растений (до 50%). Подвижные генетические элементы обычно рассеяны по геному, но могут концентрироваться в отдельных участках хромосом.
Виды мобильных элементов эукариот
Различают два (по признаку молекулярных механизмов перемещения) основных класса подвижных генетических элементов:
Похожие работы
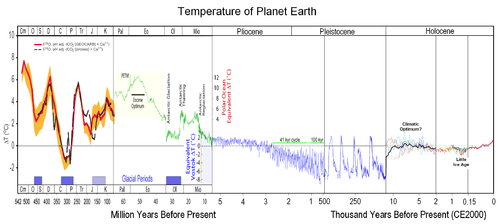
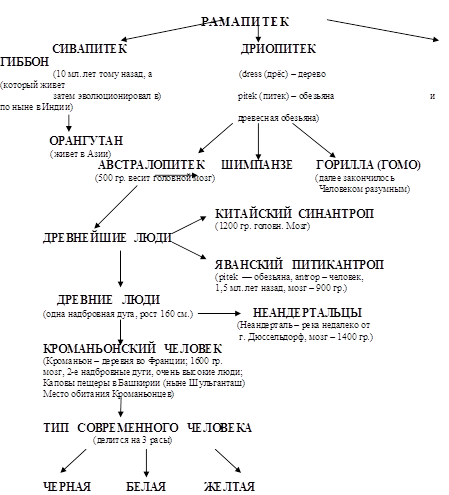

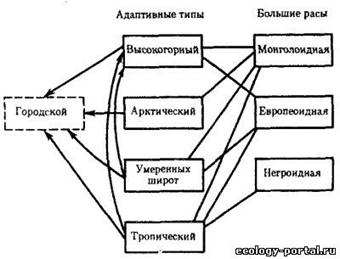

... важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии. Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную (" ...
... в области геронтологии. Это общая теория старения. Само собой разумеется, речь идет только об эскизах. Собственно это еще не общая теория старения, а только её очерк. Благодаря тому, что и как я сделал, моя скромная работа весьма отличается от известных до сих пор трудов по геронтологии. К сожалению, я не могу сразу предложить точные дефиниции, но, по крайней мере, имею возможность сделать ...
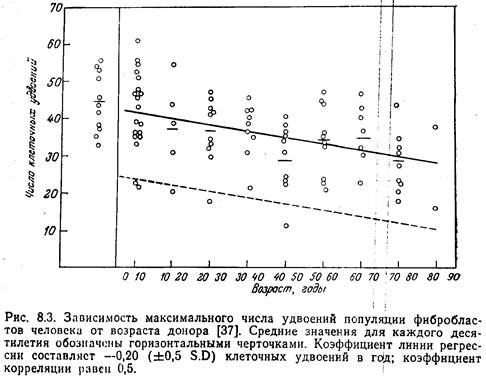
... - ядре, митохондриях, мембранах и др; в разных типах клеток- нервных, секреторных, иммунных, печёночных и др. Темп возрастных изменений определяется соотношением процессов старения и витаукта. Механизмы могут быть разделены на 2 группы. 1. Генотипические- генетически запрограммированные механизмы: а) система антиоксидантов, связывающая свободные радикалы; б) система микросомального ...

... , а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны. 3. Центральная догма и принцип Вейсмана принимаются. 4. Эволюция осуществляется путем изменения частот генов. 5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и ...
0 комментариев