Навигация
1. Транспозоны
Эти элементы ограничены инвертированными повторами (последовательностями, направленными навстречу друг другу), как и некоторые транспозоны прокариот. Примерами их могут служить Р-элемент дрозофилы и Ас-элемент кукурузы (рис. 92 Коничев). В геномах этих организмов насчитывается по 30 – 50 копий таких элементов. Полные копии (часть копий дефектна, так как имеет внутренние делеции) содержат открытые рамки считывания, кодирующие транспозазу (рис. 92).
Схема перемещения транспозона показана на рис. 93. Инвертированные повторы необходимы для перемещения элемента, которое осуществляется благодаря их сближению друг с другом и узнаванию транспозазами. Инвертированные повторы сближаются и точно отрезаются от соседних участков ДНК хозяина. Вырезанный транспозон внедряется в район вносимого транспозазой разрыва в молекуле-мишени и сшивается с ДНК хозяина в новом месте. Разрыв и зашивание осуществляются транспозазой и вспомогательными белками. Транспозаза может кодироваться как самим подвижным элементом, который будет перемещаться, так и другой копией элемента, локализованной в том же геноме в отдалении. Р-элементы обычно встраиваются в определенные сайты, консенсусная последовательность которых такова: 5'-ГГЦЦАГАГ При встраивании Р-элемента этот сайт-мишень дуплицируется.
![]()
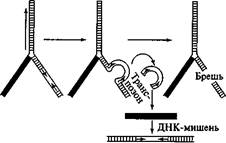
Встраивание
Рис. 93 Перемещение транспозона
Брешь в ДНК после вырезания транспозона может заделываться либо с помощью гомологичного отрезка ДНК, либо за счет сшивания разорванных концов (это особенно характерно для транспозонов растений).
Эффект от перемещения транспозона эукариот
Р-элемент содержится только в некоторых линиях D. melanogaster. Скрещивание самок без Р-элемента с самцами, несущими Р-элементы, приводит у гибридов к транспозициям Р-элемента, которые наблюдаются только в клетках зародышевого пути. Совокупность наблюдаемых эффектов называют гибридным дисгенезом. Дисгенные мухи часто оказываются стерильными, а если они фертильны, то у их потомков обнаруживается множество мутаций и эти признаки передаются следующим поколениям. Поэтому линии с Р-элементом и без него выглядят как репродуктивно изолированные, по крайней мере частично. Биологическая изоляция играет огромную роль в процессе эволюции. Изоляция линий вызвана активацией транспозиций Р-элемента, присутствующего в одной из них. Причина, почему транспозиции Р-элемента ограничены зародышевыми клетками, объясняется тем, что только в клетках-предшественниках гамет осуществляется такой тип сплайсинга, который приводит к образованию непрерывной открытой рамки трансляции, кодирующей транспозазу (см. рис. 92). Ограничение транспозиции зародышевыми клетками имеет определенный смысл, поскольку обеспечивает выживание особей, несущих гаметы, в которых произошли геномные перестройки вследствие транспозиции Р-элемента. Подобный «геномный шок», сопровождающийся высокой частотой мутагенеза, может обеспечить большую степень геномной изменчивости, которая послужит материалом для отбора в процессе эволюции.
Перемещения Ас-элемента происходят и в соматических клетках кукурузы. За ними можно следить по распределению мутантных участков ткани, например лишенных пигмента вследствие инактивации гена, определяющего пигментацию. Потомство клетки, содержащей только инактивированные гены, также будет лишено пигмента. Вырезание мобильного элемента приводит к реактивации гена. В связи с этим был сделан вывод о регуляторной функции перемещающихся элементов и они были названы контролирующими (Б. Мак-Клинток).
В целом подвижные элементы эукариот представляют собой чрезвычайно разнородную популяцию. Существует мнение о том, что они не влияют на фенотип организма и размножаются в геноме лишь благодаря особенностям своей структуры, в результате чего постепенно заселяют геном. Предполагается, что они составляют часть так называемой эгоистичной (см. с. 156) ДНК, размножение которой ограничивается естественным отбором. Отбор устраняет те случаи, когда внедрение эгоистичной ДНК сопровождается вредными последствиями. На определенных стаднях эволюции эти элементы могут использоваться для целей, которые в дальнейшем обеспечат селективные преимущества организму.
2. Ретротранспозоны.
А) с длинными концевыми повторами
– Сходны по своей структуре с проретровирусами, которые внедряются в геном, используя механизмы обратной транскрипции.
– Эти элементы содержат «тело» размером 5–8 тыс. н.п., ограниченное прямыми длинными концевыми повторами (ДКП, или LTR – от англ. 1опg terminal repeats), обычно содержащими по 300–400 н. п. (рис. 94).
– Число копий этих элементов, принадлежащих к одному семейству, достаточно постоянно для вида, но варьирует от нескольких копий до сотен тысяч копий в зависимости от типа ретротранспозона.
– В составе «тела» элемента обнаруживаются открытые рамки считывания для обратной транскриптазы и нуклеазы (интегразы).
– Разные копии одного семейства могут иметь вставки и делеции как в ДКП, так и в самом элементе.
– По флангам ДКП находятся короткие прямые повторы.
Способ перемещения ретротранспозонов с длинными концевыми повторами
– предполагает его транскрипцию с помощью РНК-полимеразы П.
– В составе ДКП имеются сайты инициации транскрипции и сигналы полиаденилирования. ДКП могут служить активными промоторами.
– Транскрипция начинается в одном ДКП (условно левом, 5'-ДКП), а заканчивается в другом (правом, 3'-ДКП).
– Синтезируемая молекула РНК транслируется с образованием ферментов, необходимых для синтеза ДНК и внедрения ее в геном.
Эта схема полностью повторяет схему образования и интеграции провируса (рис. 95). Остается открытым вопрос: произошли ли ретровирусы из ретротранспозонов или, напротив, ретротранспозоны возникли из вирусов в результате потери способности к заражению? Подавляющая часть ретротранспозонов (при сравнении их с ретровирусами) либо потеряла ген оболочки вируса, либо еще не приобрела его. Таким образом, ретротранспозоны представляют собой внутригеномные, неинфекционные элементы, способные лишь к самовоспроизведению и «подзаражению» того же генома.
Внедрение ретротранспозона внутрь гена или около гена вызывает разные эффекты:
– происходит инактивация гена (рис. 96 а).
– при интеграции в район промотора на 5'-фланге гена ретротранспозон может резко активировать экспрессию гена, обеспечивая транскрипцию с собственного промотора (рис. 96 б1). Если в этом случае подвижный элемент оказался около протоонкогена, то результатом может быть сверхпродукция белка и злокачественное перерождение клетки.
– это же возможно при воздействии энхансеров, привносимых мобильным элементом (рис. 96 б2).
– Особые возможности для перенесения и приобретения регуляторных сигналов возникают тогда, когда элемент (ретротранспозон) удаляется за счет гомологичной комбинации между ДКП с идентичными повторами (см. (рис. 96 в). В результате сохраняется лишь один ДКП на месте внедрения ретротранспозона.
Представленные случаи внедрения элемента сопровождаются мутациями с самыми разными фенотипическими проявлениями обусловленными подавлением образования или, наоборот, гиперпродукцией белка.
Перемещение элементов по геному способствует
– распространению регуляторных сигналов (сайтов инициации транскрипции, сигналов полиаденилирования, или энхансеров), что делает значительной роль мобильных элементов в эволюции систем регуляции.
Б) Ретротранспозоны, которые не несут длинных концевых повторов. Механизм внедрения их
– несколько иной,
– но также осуществляется с помощью обратной транскрипции.
– К ним относятся представители семейства L1 населяющие геном человека.
– Репликация элемента без ДКП сопряжена непосредственно с районом будущего внедрения ретротранспозона (рис. 97).
– РНК, образовавшаяся при транскрипции элемента, перемещается к достаточно случайному месту разрыва ДНК-мишени,
– часто сшивается с одной из нитей ДНК.
– Сюда устремляются и необходимые для интеграции белки – обратная транскриптаза и интеграза.
– Другая, комплементарная нить ДНК служит затравкой для копирования РНК-копии элемента с участием обратной транскриптазы.
– Фермент копирует сначала небольшой участок ДНК-мишени, а затем меняет матрицу и копирует РНК,
– после чего РНК удаляется и образуется вторая комплементарная нить ДНК.
Ретротранспозоны без ДКП участвуют в сохранении концов хромосом в ряду поколений.
Подобную функцию у многих организмов выполняет теломераза. У ряда насекомых, в том числе и у дрозофилы, отсутствует теломеразная активность. Концы ДНК у них удлиняются за счет перемещений ретротранспозонов и в этом случае выступают как компоненты генома, спасающие хромосому от укорачивания. Ретротранспозоны перемещаются, образуя повторяющуюся структуру, в которой элементы соединены друг с другом по типу «голова к хвосту». Сначала на РНК-транскрипте как на матрице с помощью обратной транскрипции строится комплементарная нить ДНК, а затем после удаления РНК-матрицы достраивается другая. Таким образом, геном хозяина приспособил эти элементы для сохранения концевых участков хромосом, для спасения хромосомы от потери генов.
Похожие работы


... важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии. Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную (" ...
... в области геронтологии. Это общая теория старения. Само собой разумеется, речь идет только об эскизах. Собственно это еще не общая теория старения, а только её очерк. Благодаря тому, что и как я сделал, моя скромная работа весьма отличается от известных до сих пор трудов по геронтологии. К сожалению, я не могу сразу предложить точные дефиниции, но, по крайней мере, имею возможность сделать ...
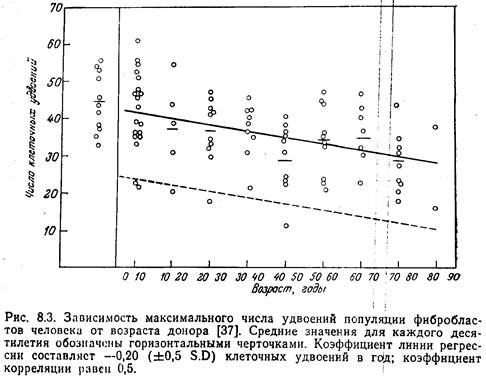
... - ядре, митохондриях, мембранах и др; в разных типах клеток- нервных, секреторных, иммунных, печёночных и др. Темп возрастных изменений определяется соотношением процессов старения и витаукта. Механизмы могут быть разделены на 2 группы. 1. Генотипические- генетически запрограммированные механизмы: а) система антиоксидантов, связывающая свободные радикалы; б) система микросомального ...

... , а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны. 3. Центральная догма и принцип Вейсмана принимаются. 4. Эволюция осуществляется путем изменения частот генов. 5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и ...
0 комментариев