Навигация
Экологические группы растений по отношению к температуре
63762
знака
0
таблиц
0
изображений
ТЕМА КУРСОВОЙ. ЭКОЛОГИЧЕСКИЕ ГРУППЫ РАСТЕНИЙ ПО ОТНОШЕНИЮ К ТЕМПЕРАТУРЕ
Содержание
Содержание
Введение 2
ГЛАВА 1. Температура как экологический фактор 4
ГЛАВА 2. Температура растений 6
ГЛАВА 3. Действие температурного стресса 11
3.1. Картина повреждения 11
3.2 .Причины гибели при перегреве 12
3.3. Гибель от охлаждения и от мороза 12
3.4. Термоустойчивость 13
3.5. Устойчивость протоплазмы 14
ГЛАВА 4. Растения и высокая температура 17
4.1. Жароустойчивость 22
4.2. Нежаростойкие виды 22
4.3. Жаровыносливые эукариоты 23
4.4. Жароустойчивые прокариоты 23
ГЛАВА 5. Влияние холода на растения и приспособления к нему 24
5.1. Нехолодостойкие растения 35
5.2. Неморозостойкие растения 35
5.3. Льдоустойчивые растения 35
Заключение 36
Литература 37
ВВЕДЕНИЕ
Приспособленность онтогенеза растений к условиям среды является результатом их эволюционного развития (изменчивости, наследственности, отбора). На протяжении филогенеза каждого вида растений в процессе эволюции выработались определенные потребности индивидуума к условиям существования и приспособленность к занимаемой им экологической нише. Жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в ходе эволюции в результате длительного действия соответствующих условий. Так, теплолюбивые растения и растения короткого дня характерны для южных широт, менее требовательные к теплу и растения длинного дня - для северных.
В природе в одном географическом регионе каждый вид растений занимает экологическую нишу, соответствующую его биологическим особенностям. Наследственность растений формируется под влиянием определенных условий внешней среды. Важное значение имеют и внешние условия онтогенеза растений.
В большинстве случаев растения и посевы (посадки) сельскохозяйственных культур, испытывая действие тех или иных неблагоприятных факторов, проявляют устойчивость к ним как результат приспособления к условиям существования, сложившимся исторически, что отмечал еще К. А. Тимирязев.
Все физиологические и биохимические процессы идут лишь в определенных температурных границах, которые обычно лежат в довольно узких пределах. Фактор тепла имеет большое значение и в географическом распределении растений. Составляя существенную часть климатических условий, он тем самым определяет северные и южные границы ареалов, зональную структуру растительного покрова.
Цель: выявить основные типы отношений растений к температуре, способы адаптаций
Задачи: 1 охарактеризовать тепловые условия местообитаний
2 охарактеризовать собственную температуру растения
3 определить последствия температурного стресса
4 рассмотреть различные виды адаптаций к высоким и низким температурам
ГЛАВА 1. ТЕМПЕРАТУРА КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
Растения — пойкилотермные организмы, т. е. их собственная температура уравнивается с температурой окружающей их среды. Однако это соответствие неполное. Конечно, тепло, выделяемое при дыхании и используемое при синтезах, вряд ли играет какую-либо экологическую роль, но все же температура надземных частей растения может значительно отличаться от температуры воздуха в результате энергообмена с окружающей средой. Благодаря этому, например, растения Арктики и высокогорий, которые заселяют места, защищенные от ветра, или растут вплотную к почве, имеют более благоприятный тепловой режим и могут достаточно активно поддерживать обмен веществ и рост, несмотря на постоянно низкие температуры воздуха. Не только отдельные растения и их части, но и целые фитоценозы обнаруживают иногда характерные отклонения от температуры воздуха. В один жаркий летний день в Центральной Европе температура на поверхности крон в лесах была на 4°С, а лугов — на 6 °С выше температуры воздуха и на 8 °С (лес) или 6 °С (луг) ниже, чем температура поверхности почвы, лишенной растительности.
Чтобы охарактеризовать тепловые условия местообитания растений, необходимо знать закономерности распределения тепла в пространстве и его динамику во времени как в отношении общеклиматических характеристик, так и конкретных условий произрастания растений.
Общее представление об обеспеченности того или иного района теплом дают такие общеклиматические показатели, как среднегодовая температура для данной местности, абсолютный максимум и абсолютный минимум (т. е. наиболее высокая и наиболее низкая температура, зарегистрированная в этом районе), средняя температура самого теплого месяца (на большей части северного полушария это июль, южного полушария - январь, на островах и в прибрежных районах — август и февраль); средняя температура самого холодного месяца (в континентальных областях северного полушария - январь, южного — июль, в прибрежных районах - февраль и август).
Для характеристики тепловых условий жизни растений важно знать не только общее количество тепла, но и его распределение во времени, от которого зависят возможности вегетационного периода. Годовую динамику тепла хорошо отражает ход среднемесячных (или среднесуточных) температур, неодинаковый на разных широтах и при разных типах климата, а также динамика максимальных и минимальных температур. Границы вегетационного сезона определяются продолжительностью безморозного периода, частотой и степенью вероятности весенних и осенних заморозков. Естественно, порог вегетации не может быть одинаковым для растений с разным отношением к теплу; для холодостойких культурных видов условно принимают 5°С, для большинства культур умеренной зоны 10°С, для теплолюбивых 15°С. Считают, что для естественной растительности умеренных широт пороговая температура начала весенних явлений составляет 5°С.
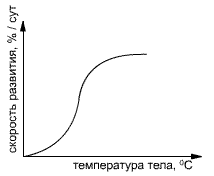
В общих чертах скорость сезонного развития пропорциональна накопленной сумме температур (стоит, например, сравнить медленное развитие растений в холодную и затяжную весну или «взрывное» начало весны при сильной волне тепла). От этой общей закономерности есть ряд отступлений: так, например, слишком высокие суммы температур уже не ускоряют, а тормозят развитие.
ГЛАВА 2. ТЕМПЕРАТУРА РАСТЕНИЙ
Наряду с тепловыми характеристиками окружающей среды необходимо знать температуру самих растений и ее изменения, поскольку именно она представляет истинный температурный фон для физиологических процессов. Температуру растений измеряют с помощью электротермометров, имеющих миниатюрные полупроводниковые датчики. Чтобы датчик не повлиял на температуру измеряемого органа, необходимо, чтобы его масса была во много раз меньше массы органа. Датчик должен быть также малоинерционным и быстро реагировать на изменения температуры. Иногда для этой цели используют термопары. Датчики или прикладывают к поверхности растения, или «вживляют» в стебли, листья, под кору (например, для измерения температуры камбия). Одновременно обязательно измеряют температуру окружающего воздуха (затенив датчик).
Температура растений весьма непостоянна. Из-за турбулентных потоков и непрерывных изменений температуры воздуха, непосредственно окружающего лист, действия ветра и т. д. температура растения варьирует с размахом в несколько десятых долей или даже целых градусов и с частотой в несколько секунд. Поэтому под «температурой растений» следует понимать более или менее обобщенную и в достаточной мере условную величину, характеризующую общий уровень нагрева. Растения как пойкилотермные организмы не имеют собственной стабильной температуры тела. Их температура определяется тепловым балансом, т. е. соотношением поглощения и отдачи энергии. Эти величины зависят от многих свойств как окружающей среды (размеры прихода радиации, температура окружающего воздуха и его движение), так и самих растений (окраска и другие оптические свойства растения, величина и расположение листьев и т. д.). Первостепенную роль играет охлаждающее действие транспирации, которое препятствует очень сильным перегревам в жарких местообитаниях. Это легко показать в опытах с пустынными растениями: стоит лишь смазать вазелином ту поверхность листа, на которой расположены устьица, и лист на глазах гибнет от перегрева и ожогов.
В результате действия всех указанных причин температура растений обычно отличается (иногда довольно значительно) от температуры окружающего воздуха. При этом возможны три ситуации:
1) температура растения выше температуры окружающего воздуха («супратемпературные» растения, по терминологии О. Ланге),
2) ниже ее («субтемпературные»),
3) равна или очень близка к ней. Первая ситуация встречается довольно часто в самых разнообразных условиях. Значительное превышение температуры растения над температурой воздуха обычно наблюдается у массивных органов растений, особенно в жарких местообитаниях и при слабой транспирации. Сильно нагреваются крупные мясистые стебли кактусов, утолщенные листья молочаев, очитков, молодила, у которых испарение воды очень незначительное. Так, при температуре воздуха 40—45°С пустынные кактусы нагреваются до 55—60°С; в умеренных широтах в летние дни сочные листья растений из родов Sempervivum и Sedum нередко имеют температуру 45°С, а внутри розеток молодила — до 50°С. Таким образом, превышение температуры растения над температурой воздуха может достигать 20°С.
Сильно нагреваются солнцем различные мясистые плоды: например, спелые томаты и арбузы на 10—15°С теплее воздуха; температура красных плодов в зрелых початках аронника — Arum maculatum доходит до 50°С. Довольно заметно бывает повышение температуры внутри цветка с более или менее закрытым околоцветником, сохраняющим от рассеивания тепло, которое выделяется при дыхании. Иногда это явление может иметь существенное адаптивное значение, например, для цветков лесных эфемероидов (пролески, хохлатки и др.), ранней весной, когда температура воздуха едва превышает 0°С.
Своеобразен и температурный режим таких массивных образований, как древесные стволы. У одиночно стоящих деревьев, а также в лиственных лесах в «безлистную» фазу (весной и осенью) поверхность стволов сильно нагревается в дневные часы, причем в наибольшей степени с южной стороны; температура камбия здесь может быть на 10—20°С выше, чем на северной стороне, где она имеет температуру окружающего воздуха. В жаркие дни температура темных стволов ели повышается до 50—55°С, что может принести к ожогам камбия. Показания тонких термопар, вживленных под кору, позволили установить, что стволы древесных пород защищены по-разному: у березы температура камбия быстрее меняется в соответствии с колебаниями температуры наружного воздуха, в то время как у сосны она более постоянна благодаря лучшим теплозащитным свойствам коры. Нагревание стволов деревьев и безлистном весеннем лесу существенно влияет на микроклимат лесного сообщества, поскольку стволы — хорошие аккумуляторы тепла.
Превышение температуры растений над температурой воздуха встречается не только в сильно прогреваемых, но и в более холодных местообитаниях. Этому способствует темная окраска или иные оптические свойства растений, увеличивающие поглощение солнечной радиации, а также анатомо-морфологические особенности, способствующие снижению транспирации. Довольно заметно могут нагреваться арктические растения: один пример - карликовая ива — Salix arctica на Аляске, у которой днем листья теплее воздуха на 2—11°С и даже в ночные часы полярного «круглосуточного дня» — на 1—3°С. Ранневесенним эфемероидам «подснежникам» нагревание листьев обеспечивает возможность достаточно интенсивного фотосинтеза в солнечные, но еще холодные весенние дни. Даже под снегом (точнее под тонким слоем полупрозрачного фирна) темноокрашенные части зимующих альпийских и арктических растений нагреваются солнечными лучами. Это приводит к образованию полостей и «парничков» вокруг растений, к более быстрому растаиванию снежной корки над ними. Когда над поверхностью снега в высокогорьях Альп и Карпат появляются темно-сиреневые колокольчатые цветки сольданелл — Soldanella alpina, S. hungarica, создается впечатление, что растения пробивают снег, «растапливая его теплотой дыхания» (мнение, распространенное в старой научно-популярной литературе). На самом деле расчеты показывают, что эта теплота ничтожно мала. Еще один интересный пример нагревания под снегом: в летнее время в Антарктиде температура лишайников бывает выше 0°С даже под слоем снега более 30 см. Очевидно, в столь суровых условиях естественный отбор сохранил формы с наиболее темной окраской, у которых благодаря такому нагреванию возможен положительный баланс углекислотного газообмена.
Довольно значительно могут нагреваться солнечными лучами иглы хвойных древесных пород зимой: даже при отрицательных температурах возможно превышение над температурой воздуха на 9—12°С, что создает благоприятные возможности для зимнего фотосинтеза. Экспериментально было показано, что если для растений создать сильный поток радиации, то даже при низкой температуре порядка —5, —6°С листья могут нагреться до 17—19°С, т. е. фотосинтезировать при вполне «летних» температурах.
Для холодных местообитаний или сезонных экологических ниш повышение температуры растения экологически очень важно, так как физиологические процессы при этом получают независимость в известных пределах от окружающего теплового фона.
Снижение температуры растений по сравнению с окружающим воздухом чаще всего отмечается в сильно освещенных и прогреваемых местообитаниях (степях, пустынях), где листовая поверхность растений сильно редуцирована (см. ниже), а усиленная транспирация способствует удалению избытка тепла и предотвращает перегрев (напомним, что для испарения 1 г воды при 20°С требуется 2438 Дж — 582 кал). Недаром иногда говорят о «гидротерморегуляции» растений. У интенсивно транспирирующих видов охлаждение листьев (разность с температурой воздуха) достигает 15°С. Это крайний пример, но и снижение на 3—4°С может предохранить от губительного перегрева.
В самых общих чертах можно сказать, что в жарких местообитаниях температура надземных частей растений ниже, а в холодных— выше температуры воздуха. Эта закономерность прослеживается и на одних и тех же видах: так, в холодном поясе гор Северной Америки, на высотах 3000—3500 м, растения теплее, а в низкогорном — холоднее воздуха.
Совпадение температуры растений с температурой окружающего воздуха встречается гораздо реже в условиях, исключающих сильный приток радиации и интенсивную транспирацию, например у травянистых растений под пологом тенистых лесов (но не на солнечных бликах), а на открытых местообитаниях — в пасмурную погоду или при дожде.
В целом, по мнению многих авторов, совпадение температуры растения и среды является исключением, а несовпадение — правилом, в связи с чем иногда говорят — с большой долей условности даже о «собственном микроклимате растений».
Различают разные экологические типы растений по отношению к температуре. У растений термофильных, или мегатермных (теплолюбивых), оптимум лежит в области повышенных температур. Они обитают в областях тропического и субтропического климата, а в умеренных поясах — в сильнопрогреваемых местообитаниях. Для криофильных,или микротермных (холодолюбивых), растений оптимальны низкие температуры. К ним принадлежат виды, живущие в полярных и высокогорных областях или занимающие холодные экологические ниши. Иногда выделяют промежуточную группу мезотермных растений.
ГЛАВА 3. ДЕЙСТВИЕ ТЕМПЕРАТУРНОГО СТРЕССА
Жара и мороз вредят жизненным функциям и ограничивают распространение вида в зависимости от их интенсивности, продолжительности и периодичности, но прежде всего от состояния активности и степени закалки растений. Стресс-—это всегда необычная нагрузка, которая не обязательно должна быть опасной для жизни, но которая непременно вызывает в организме «реакцию тревоги», если только он не находится в выраженном состоянии оцепенения. Покоящиеся стадии, такие, как сухие споры, а также пойкилогидрические растения в высохшем состоянии, нечувствительны, так что они могут пережить без повреждения любую отмеченную на Земле температуру.
Протоплазма вначале отвечает на стресс резким усилением метаболизма. Повышение интенсивности дыхания, которое наблюдается в качестве стрессовой реакции отражает попытку исправить уже появившиеся дефекты и создать ультраструктурные предпосылки для приспособления к новой ситуации. Стрессовая реакция — это борьба механизмов адаптации, с деструктивными процессами в протоплазме, ведущими к ее гибели.
Гибель клеток от перегрева и холода. Если температура переходит критическую точку, клеточные структуры и функции могут повреждаться так внезапно, что протоплазма тотчас же отмирает. В природе такое внезапное разрушение нередко происходит при эпизодических морозах, например при поздних заморозках весной. Но повреждения могут возникать и постепенно; отдельные жизненные функции выводятся из равновесия и угнетаются, пока, наконец, клетка не отомрет в результате прекращения жизненноважных процессов.
Похожие работы
... которой создается более теплый микроклимат; у обитателей тундры возникают стелящиеся, или шпалерные, и карликовые формы. По отношению к теплу выделяют следующие экологические группы: Эвритермные и стенотермные организмы (см. предыдущий урок) Термофилы и криофилы (теплолюбивые и холодолюбивые) По степени адаптации к условиям дефицита тепла различают нехолодостойкие (гибнут при температуре ...
... , опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, ...
... сумеречного освещения в водных глубинах и пещерах. В разных местообитаниях различаются не только интенсивность света, но и его спектральный состав, продолжительность освещения, пространственное и временное распределение света разной интенсивности и т. д. Соответственно, разнообразны и приспособления растений к жизни при том или ином световом режиме. Экологические группы растений по отношению к ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев