Навигация
3.1. Картина повреждения
Различные жизненные процессы неодинаково чувствительны к температуре. Сначала прекращается движение протоплазмы, интенсивность которого непосредственно зависит от энергоснабжения за счет процессов дыхания и от наличия высокоэнергетических фосфатов. Затем снижаются фотосинтез и дыхание. Для фотосинтеза особенно опасна жара, дыхание же наиболее чувствительно к холоду. У поврежденных холодом или жарой растений после возвращения в умеренные условия уровень дыхания сильно колеблется и часто бывает ненормально повышен. Повреждение хлоропластов ведет к длительному или необратимому угнетению фотосинтеза. В конечной стадии утрачивается полупроницаемость биомембран, разрушаются клеточные компартменты, особенно тилакоиды пластид, и клеточный сок выходит в межклетники..
3.2. Причины гибели при перегреве
Высокая температура быстро приводит к гибели из-за повреждения мембран и прежде всего в результате инактивации и денатурации белков. Даже если из строя выходят только немногие, особо термолабильные ферменты, это ведет к расстройству обмена нуклеиновых кислот и белков и в конце концов — тоже к гибели клеток. Растворимые азотистые соединения накапливаются при этом в таких больших концентрациях, что они диффундируют из клеток и теряются; кроме того, образуются ядовитые продукты распада, которые не могут больше обезвреживаться в ходе обмена веществ.
3.3. Гибель от охлаждения и от мороза
При повреждении протоплазмы холодом следует различать, вызвано ли оно самой по себе низкой температурой или же замерзанием. Некоторые растения тропического происхождения повреждаются уже при снижении температуры до нескольких градусов выше нуля. Подобно гибели от перегрева, гибель от охлаждения тоже бывает прежде всего связана с дезорганизацией обмена нуклеиновых кислот и белков, но здесь играют роль также нарушения, проницаемости и прекращение тока ассимилятов.
Растения, которым охлаждение до температур выше нуля не причиняет вреда, повреждаются только при температурах ниже нуля, т. е. в результате образования льда в тканях. Богатые водой, незакаленные протопласты могут легко замерзать; при этом внутри клетки мгновенно образуются ледяные кристаллики, и клетка погибает. Чаще всего лед образуется не в протопластах, а в межклетниках и клеточных стенках. Такое образование льда называют внеклеточным. Выкристаллизовавшийся лед действует как сухой воздух, так как упругость пара надо льдом ниже, чем над переохлажденным раствором. В результате от протопластов отнимается вода, они сильно сжимаются (на 2/з своего объема) и концентрация растворенных веществ в них возрастает. Перемещение воды и замерзание продолжаются до тех пор, пока в протоплазме не установится равновесие сосущих сил между льдом и водой. Положение равновесия зависит от температуры; при температуре —5°С равновесие наступает примерно при; 60 бар, а при —10°С — уже при 120 -бар. Таким образом, низкие температуры действуют на протоплазму так же, как высыхание. Морозоустойчивость клетки более высока, если вода прочно связана со структурами протоплазмы и осмотически связана. При обезвоживании цитоплазмы (безразлично, в результате засухи или замерзания) инактивируются ферментные системы, ассоциированные с мембранами, - системы, участвующие главным образом в синтезе АТФ и в процессах фосфорилирования (Хебер, Сантариус, 1979). Инактивацию, вызывают чрезмерные и потому токсичные концентрации ионов. солей и органических кислот в незамерзшем остаточном растворе. Напротив, сахара, производные сахаров, определенные аминокислоты и белки защищают биомембраны и ферменты от вредных веществ (Максимов, Туманов, Красавцев, 1952). Наряду с этим есть указания на то, что при замерзании белки денатурируются, что также ведет к повреждению мембран (Левитт 1980).
3.4. Термоустойчивость
Термоустойчивость — это способность организма переносить большую жару или холод без необратимого повреждения. Термоустойчивость растения складывается из способности протоплазмы переносить экстремальные температуры (толерантность по Дж. Левитту) и из эффективности мер, которые замедляют или предотвращают развитие повреждений (избегание).
Меры, позволяющие избегать повреждения
Возможные способы защиты клеток от температурного повреждения немногочисленны и не очень эффективны. Изоляция от перегрева и охлаждения может дать лишь кратковременную защиту. Так. например, в густых кронах деревьев или у подушечных растений почки листья и цветки, находящиеся в глубине и ближе к земле, меньше подвергаются опасности замерзнуть в результате отдачи тепла путем излучения, чем наружные части растения. Виды хвойных с особенно толстой коркой лучше выдерживают пожары в подлеске. Общее же значение имеют главным образом две защитные меры: замедление образования льда в тканях и (при жаре) охлаждение путем отражения падающих лучей и с помощью транспирации.
3.5. Устойчивость протоплазмы
Длительное и регулярно повторяющееся воздействие крайних. температур растения могут выдерживать только в том случае, если сама протоплазма жаро- или морозоустойчива. Эта особенность обусловлена генетически и поэтому у разных видов и даже сортов выражена в разной степени. Однако это не такое свойство, которое присуще растению постоянно и всегда в одинаковой мере. Проростки, весенние побеги древесных растений в период их интенсивного растяжения, культуры микроорганизмов в фазе экспоненциального роста вряд ли способны закаливаться и поэтому чрезвычайно чувствительны к температуре.
Устойчивость к образованию льда и закаливание к действию мороза. В областях с сезонным климатом наземные растения приобретают осенью «льдоустойчивость», т. е. способность переносить образование льда в тканях. Весной, с распусканием почек они снова утрачивают эту способность, и теперь замерзание приводит к их вымерзанию. Таким образом, холодостойкость многолетних растений вне тропиков регулярно колеблется в течение года между минимальной величиной в период вегетации и максимумом в зимнее время. Льдоустойчивость формируется постепенно осенью. Первая предпосылка для этого—переход растения в состояние готовности к закаливанию, наступающее только тогда, когда заканчивается рост. Если готовность к закаливанию достигнута, то процесс закаливания может начинаться. Этот процесс состоит из нескольких фаз, каждая из которых подготавливает переход к следующей. По теории, разработанной И.И. Тумановым, закаливание к морозу, у озимых злаков и плодовых; деревьев (эти растения изучались наиболее основательно) начинается многодневным (до нескольких недель) воздействием температур чуть выше нуля. На этой фазе, предшествующей закаливанию, в протоплазме накапливаются сахара и другие защитные вещества, клетки становятся беднее водой, а центральная вакуоль распадается на множество мелких вакуолей. Благодаря этому протоплазма оказывается подготовленной к следующей фазе, проходящей при регулярных слабых морозах от —3 до—5°С. При этом ультраструктуры и ферменты протоплазмы перестраиваются таким образом, что клетки переносят обезвоживание, связанное с образованием льда. Только после этого растения могут, не подвергаясь, опасности, вступать в заключительную фазу процесса; закаливания, которая при непрерывном морозе по меньшей мере от —10 до-—15 °С делает протоплазму в высшей степени морозоустойчивой.
Зоны эффективных температур для разных видов различны . Готовые к закаливанию сеянцы березы, которые до начала процесса закаливания вымерзали бы при температуре от —15-до —20°С, переносят после окончания первой фазы закаливания; уже —35 °С, а при полной закалке они выдерживают даже охлаждение до —195°С. Таким образом, холод сам по себе стимулирует процесс закаливания. Если мороз ослабевает, то протоплазма опять переходит в первую фазу закаливания, однако устойчивость, снова может быть поднята холодными периодами до наивысшего уровня, пока растения остаются в состоянии покоя.
.В зимний период на сезонный ход морозостойкости накладываются кратковременные (индуцированные), адаптации, благодаря которым уровень устойчивости быстро приспосабливается к изменениям погоды. Холод больше всего способствует закаливанию в начале зимы. В это время устойчивость может за несколько дней подняться до наивысшего уровня. Оттепель, особенно в конце зимы, вызывает быстрое снижение устойчивости растений, но в середине зимы после выдерживания в течение .нескольких дней при температуре от +10 до +20 °С растения в значительной мере теряют закалку. Способность изменять морозоустойчивость под влиянием холода и тепла, т. е. диапазон индуцируемых адаптации устойчивости, является конституционным признаком отдельных видов растений.
После окончания зимнего покоя способность к закаливанию и вместе с тем высокая степень закалки быстро утрачиваются. Весной существует тесная связь между активированием распускания почек и ходом изменения устойчивости.
ГЛАВА 4. РАСТЕНИЯ И ВЫСОКАЯ ТЕМПЕРАТУРА
Как уже указывалось, в открытых местообитаниях с сильной инсоляцией и высокими температурами надземные части растений (особенно слабо транспирирующих) могут нагреваться до 45 - 60°С. Нагревание напочвенных и наскальных лишайников достигает 60—65°С (иногда в течение довольно продолжительного времени). «Полюс жары» в растительном мире занимают растения термальных (горячих) источников Камчатки, Исландии, Йеллоустоунского парка в США и др. Наивысшая температура, при которой найдены живые сине-зеленые водоросли, 85°С, бактерии — 88°С. Высшие растения в термальных водах отсутствуют (лишь один вид ряски живет при 32—35°С). Интересно, что обитатели термальных вод живут при температурах, очень близких к летальному пределу температур: водоросль Oscillatoria, живущая на о. Ява в воде с температурой 64°С, погибает при 68°С уже через 5—10 мин! Водоросли выдерживают очень высокие температуры (67—75°С) только в условиях полного солнечного освещения, а в затененных источниках не выносят и более низкой температуры порядка 50—55°С.
Действие экстремальных высоких температур влечет за собой целый ряд опасностей для растений: сильное обезвоживание и иссушение, ожоги, разрушение хлорофилла, необратимые расстройства дыхания и других физиологических процессов, наконец, тепловую денатурацию белков, коагуляцию цитоплазмы и гибель. Перегрев почвы приводит к повреждению и отмиранию поверхностно расположенных корней, к ожогам корневой шейки.
В защитных приспособлениях растений к высоким температурам использованы разные пути адаптации. Анатомо-морфологические черты, предотвращающие перегрев, в основном те же самые, что служат растению для ослабления прихода радиации к тканям надземных частей. Это густое опущение, придающее листьям светлую окраску и усиливающее их способность к отражению; блестящая поверхность; уменьшение поверхности, поглощающей радиацию, — вертикальное и меридиональное положение листьев; свертывание листовых пластинок у злаков; общая редукция листовой поверхности и т. д. Эти же особенности строения одновременно способствуют уменьшению потери воды растением. Таким образом, комплексное действие экологических факторов на растение находит отражение и в комплексном характере адаптации. Поэтому трудно отличить те черты структуры, которые служили бы растению только «тепловой защитой» за редкими исключениями (например, развитие пробковой ткани или воздухоносной ткани близ корневой шейки у некоторых пустынных растений).
Весьма действенной физиологической адаптацией к перегреву служит усиленная транспирация, роль которой в терморегуляции растений уже подчеркивалась выше. Ряд авторов придает значение высокому содержанию у жаростойких растений защитных веществ (слизи, органические кислоты и др.). В адаптации растений к высоким температурам принимают участие весьма тонкие механизмы на клеточном и субклеточном уровне, например, сдвиги температурного оптимума активности важнейших ферментов. По современным представлениям (Александров, 1975), в основе устойчивости организмов к действию высоких температур (как и других экстремальных воздействий) лежит особое свойство структуры белковых молекул — сочетание прочности и гибкости, позволяющее им поддерживать структуру и функциональную активность в крайних условиях.
Своеобразное физиологическое приспособление к температуре среды, превышающей адаптивные возможности растений, — переход в состояние анабиоза, которое в этих случаях особенно часто встречается среди низших растений.
Наконец, следует упомянуть еще об одном способе адаптации растений к чрезмерно высоким температурам — это занятие ими временных экологических ниш (или микрониш), защищенных от сильной инсоляции и перегрева. У некоторых растений вся вегетация сдвигается на сезон с более благоприятными тепловыми условиями. Так, в растительном покрове пустынь и степей есть группа видов, начинающая вегетацию очень рано весной и успевающая ее закончить еще до наступления летней жары и сухости. Они переживают эти условия в состоянии летнего покоя в виде семян (эфемеры-однолетники: крупка, или веснянка весенняя, — Erophila verna, рогоглавник—Ceratocephalus falcatus, бурачок — Alyssum desertorum и др.) или подземных органов — луковиц, клубней, корневищ (эфемероиды-многолетники: тюльпаны, крокусы, гадючий лук, мятлик луковичный — Роа bulbosa и др.). Эта сезонная адаптация, связанная с перестройкой всего годичного цикла развития, обеспечивает растениям надежную защиту от жары даже в районах самых жарких пустынь.
Интересный пример того, как растения сильно прогреваемых местообитаний используют для активной жизнедеятельности даже небольшие прохладные отрезки времени, представляют лишайники жаркой и сухой пустыни Негев (Северная Африка), у которых фотосинтез идет только в самые первые прохладные часы после восхода солнца. Соответственно температурный оптимум фотосинтеза у них оказывается неожиданно низким для пустынных растений (около 20°С).
Экологические различия устойчивости растений к высоким температурам. Количественную характеристику жаростойкости растений дать не так просто, поскольку известно, что повреждающее действие экстремального фактора на живой организм зависит не только от интенсивности самого фактора, но и от продолжительности его влияния. Так, если растение в течение нескольких минут может выдержать 50—55°С, то при многочасовых экспозициях предельная температура окажется гораздо ниже, например, порядка 45°С. Сравнивать приводимые в литературе данные о температурной выносливости растений очень трудно, поскольку разные авторы используют в экспериментах неодинаковые по длительности экспозиции и различные критерии повреждения высокой температурой (появление видимых повреждений, начало отмирания листьев, снижение дыхания или фотосинтеза и т. д.).
При действии на клетку экстремальных высоких температур одновременно имеют место как повреждения и нарушения жизнедеятельности клетки, так и процессы адаптации и восстановления повреждений (репарации). В связи с этим различают (Александров, 1975) первичную теплоустойчивость - непосредственную реакцию клетки на повышение температуры, определяемую по различным признакам нарушения работы клетки при кратковременном (5-минутном) нагреве, и общую теплоустойчивость, определяемую при более длительных экспозициях, когда успевают включиться адаптационные и репараторные механизмы.
При сопоставлении общей теплоустойчивости у растений из различных по тепловым условиям местообитаний выявляется общая закономерность: четкое соответствие между температурными условиями обитания вида (в период активной жизни особей) и его выносливостью к высоким температурам. Можно проследить связь выносливости не только с общим температурным фоном, но и с температурным режимом листьев. Так, у африканских пустынных и саванных растений с интенсивной транспирацией, сильно охлаждающей листья, теплоустойчивость гораздо ниже, чем у видов со слабым транспирационным охлаждением. Такое же явление было обнаружено и у многих представителей средиземноморской флоры, причем различие теплоустойчивости у растений с разной интенсивностью транспирации достигало 12°С.
Экологические различия первичной теплоустойчивости также проявляются достаточно четко, причем особенно хорошо они видны при сравнении близких видов, живущих в условиях различного теплового фона. У северных форм теплоустойчивость ниже, чем у более южных, а у эфемерных и эфемероидных, вегетирующих ранней весной, ниже, чем у видов с летней вегетацией.
Есть отличия и у растений одних и тех же географических районов, но разных экологических ниш. У водорослей, живущих в литоральной (приливно-отливной) зоне и периодически подвергающихся высыханию и нагреванию на воздухе, общая устойчивость к нагреву, как и первичная теплоустойчивость, заметно выше (на 12— 13°С), чем у постоянно погруженных сублиторальных. Такие же различия теплоустойчивости в зависимости от глубины обитания и у водных цветковых растений.
Даже у одной и той же особи теплоустойчивость по-разному нагреваемых частей неодинакова: например, у сильно транспирирующего листа хлопчатника предел устойчивости 46°С, а у коробочек—50°С (последние обычно имеют температуру на несколько градусов выше, чем окружающий воздух). Интересно в этом отношении различие весенних и летних листьев медуницы Pulmonaria obscura; у первых, развивающихся в более прохладное время, теплоустойчивость заметно ниже, чем у летних.
Теплоустойчивость клеток растений - свойство динамичное, меняющееся при изменении температуры окружающей среды. Обнаружены два способа ее модификационного изменения (Александров, 1975). Один из них - так называемая тепловая настройка встречается у водорослей. Их теплоустойчивость смещается соответственно температуре среды, причем довольно быстро — в течение нескольких часов (так, у литоральной водоросли Fucus vesiculosus во время отлива теплоустойчивость повышается по сравнению с периодом прилива). Такое смещение обратимо.
Другой путь приведения теплоустойчивости клеток в соответствие с изменившимся тепловым фоном обнаружен у клеток мохообразных и цветковых растений: у них теплоустойчивость остается стабильной, если изменения температуры не выходят за пределы оптимальных и близких к оптимуму, но повышается при кратковременном действии высоких (супероптимальных) температур. Это явление было названо «тепловой закалкой». Оно наблюдается в природных условиях в периоды значительного повышения температур, благодаря чему растения способны переносить наиболее жаркие дни лета. Например, в Туркмении у ряда видов (злаков-—Aristida karelini, Arundo donax, древесных пород — Catalpa speciosa, Morus alba и др.) обнаружено, что в результате «тепловой закалки» теплоустойчивость повышается в самые жаркие летние месяцы и часы дня, так что в целом динамика первичной теплоустойчивости клеток хорошо согласуется с ходом температуры в течение не только вегетационного периода, но и в течение дня.
Сезонная динамика устойчивости растений к нагреванию — повышение ее в наиболее жаркий период года — проявляется и во многих других случаях.
Все сказанное до сих пор относилось к активным фазам жизненных циклов растений. Совершенно особое положение занимают покоящиеся стадии (семена, споры) или растения в состоянии анабиоза. Жароустойчивость резко снижается при увлажнении и тем более в начале процессов роста и развития.
Похожие работы
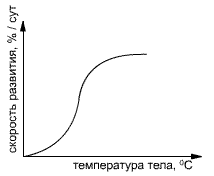
... которой создается более теплый микроклимат; у обитателей тундры возникают стелящиеся, или шпалерные, и карликовые формы. По отношению к теплу выделяют следующие экологические группы: Эвритермные и стенотермные организмы (см. предыдущий урок) Термофилы и криофилы (теплолюбивые и холодолюбивые) По степени адаптации к условиям дефицита тепла различают нехолодостойкие (гибнут при температуре ...
... , опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, ...
... сумеречного освещения в водных глубинах и пещерах. В разных местообитаниях различаются не только интенсивность света, но и его спектральный состав, продолжительность освещения, пространственное и временное распределение света разной интенсивности и т. д. Соответственно, разнообразны и приспособления растений к жизни при том или ином световом режиме. Экологические группы растений по отношению к ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев