Навигация
4.1. Жароустойчивость
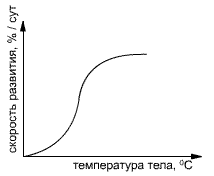
Жароустойчивость зависит от продолжительности воздействия тепла, т. е. подчиняется закону дозы: более умеренная жара при большой продолжительности оказывает такое же повреждающее действие, как и кратковременная сильная жара. Поэтому условились характеризовать устойчивость переносимостью определенных температур при их получасовом воздействии. Если бы вместо этого высокая температура поддерживалась в течение часа, то пределы устойчивости лежали бы примерно на 1—2°С ниже. По отношению к жароустойчивости можно различать следующие группы растений
4.2. Нежаростойкие виды.
В эту группу можно объединить все виды, которые повреждаются уже при 30—40°С в крайнем случае при 45 °С: эукариотические водоросли и подводные листостебельные растения, лишайники в набухшем состоянии (которые однако, при сильной инсоляции скоро высыхают и в таком состоянии вполне жаростойки) и, наконец, большинство мягколистных наземных растений, Различные фитопатогенные бактерии и вирусы также разрушаются при относительно низких температурах (например, вирус увядания томатов при 40-45°). Все эти виды могут заселять лишь такие местообитания, в которых они не подвергаются слишком большому перегреву, а более сильную жару выносят разве только в том случае, если они способны эффективно понижать собственную температуру за счет транспирации (виды “снижающие температуру", по О. Ланге).
4.3. Жаровыносливые эукариоты.
Растения солнечных и сухих местообитаний, как правило, обладают высокой способностью закаливаться по отношению к жаре; они переносят получасовое нагревание до 50—60 °С. Температура немного выше 60 °С является, очевидно, непереходимой границей для высокодифференцированных растительных клеток (с ядром и другими органеллами).
4.4. Жароустойчивые прокариоты.
Некоторые термофильные прокариоты переносят чрезвычайно высокие температуры: бактерии до 90°С, сине-зеленые водоросли до 75"С. Они обладают, так же как и термостабильные вирусы, особенно устойчивыми нуклеиновыми кислотами и белками.
Жаростойкость - очень специфичное свойство: даже близко родственные виды одного и того же рода могут заметно различаться по этому признаку. Особенно характерные различия в устойчивости, которые можно связать с условиями существования в естественных ареалах данных растений, возникли в ходе эволюции и отбора. Так, например, жаростойкость листьев многих видов в тундре составляет 42, в тайге 44 и в жарких полупустынях 47°С.
ГЛАВА 5. ВЛИЯНИЕ ХОЛОДА НА РАСТЕНИЯ И ПРИСПОСОБЛЕНИЯ К НЕМУ
О пределе холода, который способны вынести растения в естественных условиях, дают представление величины предельно низких температур на земном шаре. Там, где зарегистрирована самая низкая температура (—90°С, станция «Восток» в Антарктиде), растительность отсутствует; а в районах, где живут растения, отмечена температура —68°С (Оймякон в Якутии, область таежных лесов из лиственницы — Larix dahurica).
Растительный покров обширных территорий земного шара (умеренные и арктические области, высокогорья) ежегодно в течение нескольких месяцев подвергается действию низких температур. Кроме того, в отдельных районах и в более теплые сезоны растения могут испытывать кратковременные воздействия пониженных температур (ночные и утренние заморозки). Наконец, есть местообитания, где вся жизнь растений проходит на весьма пониженном температурном фоне (арктические снежные и морские водоросли, приснежная—нивальная растительность в высокогорьях). Неудивительно, что естественный отбор выработал у растений ряд защитных приспособлений к неблагоприятному действию холода.
Кроме непосредственного влияния низкой температуры на растения под действием холода возникают и другие неблагоприятные явления. Например, уплотнение и растрескивание замерзшей почвы приводит к разрыву и механическому повреждению корней, образование ледяной корки на поверхности почвы ухудшает аэрацию и дыхание корней. Под толстым и долго лежащим снежным покровом при температуре около 0°С наблюдается зимнее «выпревание», истощение и гибель растений в связи с расходом резервных веществ на дыхание, грибными заболеваниями («снежная плесень») и т. д., а в случае избыточно увлажненной почвы для растений опасно также зимнее «вымокание». В тундре и северной тайге распространено явление морозного «выпирания» растений, которое вызывается неравномерным замерзанием и расширением почвенной влаги. При этом возникают силы, выталкивающие растение из почвы, в результате чего происходит «выпучивание» целых дернин, оголение и обрывы корней и т. д. вплоть до повала небольших деревьев. Поэтому кроме собственно холодостойкости (или морозостойкости) — способности переносить прямое действие низких температур, различают еще зимостойкость растений - способность к перенесению всех перечисленных выше неблагоприятных зимних условий.
Особо следует остановиться на том, как влияет на растения низкая температура почвы. Холодные почвы в сочетании с умеренно-теплым режимом воздушной среды растений (а иногда и со значительным нагреванием надземных частей растений) — явление нередкое. Таковы условия жизни растений на болотах и заболоченных лугах с тяжелыми почвами, в некоторых тундровых и. высокогорных местообитаниях и в обширных областях вечной мерзлоты (около 20% всей суши), где в период вегетации оттаивает лишь неглубокий, так называемый «деятельный» слой почвы. В условиях пониженных температур почвы после снеготаяния (0—10°С) проходит значительная часть вегетации ранневесенних лесных растений — «подснежников». Наконец, кратковременные периоды резкого несоответствия холодных почв и прогретого воздуха испытывают ранней весной многие растения умеренного климата (в том числе и древесные породы).
Еще в прошлом веке немецкий физиолог Ю. Сакс показал, что при охлаждении почвы до околонулевых температур (обкладывание горшка льдом) может наступить завядание даже обильно политых растений, поскольку при низких температурах корни не способны интенсивно всасывать воду. На этом основании в экологии распространилось мнение о «физиологической сухости» местообитаний с холодными почвами (т. е. недоступности влаги растениям при ее физическом обилии). При этом упускали из виду, что Сакс и другие физиологи свои опыты производили с достаточно теплолюбивыми растениями (огурцы, тыква, салат и др.) и что в природных холодных местообитаниях растения, для которых низкие температуры почв служат естественным фоном, возможно, реагируют на них совсем иначе. Действительно, современные исследования показали, что у большинства растений тундр, болот, у ранневесенних лесных эфемероидов отсутствуют те явления угнетения (затруднение всасывания воды, расстройства водного режима и т. д.), которые могли бы быть вызваны «физиологической сухостью» холодных почв. Это же показано и для многих растений в областях вечной мерзлоты. Вместе с тем нельзя полностью отрицать угнетающее влияние низких температур на всасывание влаги и другие стороны жизнедеятельности корней (дыхание, рост и др.), а также на активность почвенной микрофлоры. Оно без сомнения имеет значение в комплексе трудных условий для жизни растений в холодных местообитаниях. «Физиологическая сухость», «физиологическая засуха» из-за низкой температуры почв возможны в жизни растений в наиболее трудных условиях, например при выращивании на холодных почвах теплолюбивых растений или ранней весной для древесных пород, когда еще необлиственные ветви сильно нагреваются (до 30—35°С) и увеличивают потерю влаги, а интенсивная работа корневых систем еще не началась.
Каких-либо специальных морфологических приспособлений, защищающих от холода, у растений нет, скорее можно говорить о защите от всего комплекса неблагоприятных условий в холодных местообитаниях, включающего сильные ветры, возможность иссушения и т. д. У растений холодных областей (или у переносящих холодные зимы) часто встречаются такие защитные морфологические особенности, как опушение почечных чешуи, зимнее засмоление почек (у хвойных), утолщенный пробковый слой, толстая кутикула, опушение листьев и т. д. Однако их защитное действие имело бы смысл лишь для сохранения собственного тепла гомеотермных организмов, для растений же эти черты, хотя и способствуют терморегуляции (уменьшение лучеиспускания), в основном важны как защита от иссушения. В растительном мире есть интересные примеры адаптации, направленных на сохранение (хотя и кратковременное) тепла в отдельных частях растения. В высокогорьях Восточной Африки и Южной Америки у гигантских «розеточных» деревьев из родов Senecio, Lobelia, Espeletia и других от частых ночных морозов существует такая защита: ночью листья розетки закрываются, защищая наиболее уязвимые части — растущие верхушки. У некоторых видов листья опушены снаружи, у других в розетке скапливается выделяемая растением вода; ночью замерзает лишь поверхностный слой, а конусы нарастания оказываются защищенными от мороза в своеобразной «ванне».
Среди морфологических адаптации растений к жизни в холодных местообитаниях важное значение имеют небольшие размеры и особые формы роста. Не только многие травянистые многолетники, но также кустарники и кустарнички полярных и высокогорных областей имеют высоту не более нескольких сантиметров, сильно сближенные междоузлия, очень мелкие листья (явление нанизма или карликовости). Кроме хорошо известного примера — карликовой березки (Betula папа), можно назвать карликовые ивы (Sahx polaris, S. arctica, S. herbacea) и многие другие. Обычно высота этих растений соответствует глубине снежного покрова, под которым зимуют растения, так как все части, выступающие над снегом, гибнут от замерзания и высыхания. Очевидно, в образовании карликовых форм в холодных местообитаниях немалую роль играют и бедность почвенного питания в результате подавления активности микробов, и торможение фотосинтеза низкими температурами. Но независимо от способа образования карликовые формы дают известное преимущество растениям в приспособлении к низким температурам: они располагаются в припочвенной экологической микронише, наиболее прогреваемой летом, а зимой хорошо защищены снежным покровом и получают дополнительный (хотя и небольшой) приток тепла из глубины почвы.
Другая адаптивная особенность формы роста — переход сравнительно крупных растений (кустарников и даже деревьев) от ортотропного (вертикального) к плагиотропному (горизонтальному) росту и образование стелющихся форм— стланцев, стлаников, стланичков. Такие формы способны образовывать кедровый стланик (Pinus pumila), можжевельник (Juniperus sibirica, J. communis, J. turkestanica), рябина и др. Ветви стланцев распластаны по земле и приподнимаются не выше обычной глубины снежного покрова. Иногда это результат отмирания ствола и разрастания нижних ветвей (например, у ели), иногда это рост дерева как бы «лежа на боку» с плагиотропным, укоренившимся во многих местах стволом и приподнимающимися ветвями (кедровый стланик). Интересная особенность некоторых древесных и кустарниковых стлаников — постоянное отмирание старой части ствола и нарастание «верхушки», в результате чего трудно определить возраст особи.
Стланики распространены в высокогорных и полярных областях, в условиях, которых уже не выдерживают древесные породы (например, на верхней границе леса). Своеобразные «стланиковые» формы в крайних условиях встречаются и у кустарничков, и даже у видов лишайников, обычно имеющих прямостоячий кустистый рост: на скалах Антарктиды они образуют стелющиеся слоевища,
В зависимости от условий возможны видоизменения роста одного и того же вида. Но есть виды, целиком перешедшие к форме стланика, например горный сосновый стланик, произрастающий в Альпах и Карпатах — Pinus mughus, выделенный в качестве самостоятельного вида из сосны горной — Pinus montana.
К числу форм роста, способствующих выживанию растений в холодных местообитаниях, принадлежит еще одна чрезвычайно своеобразная — подушковидная. Форма растения-подушки образуется в результате усиленного ветвления и крайне замедленного роста скелетных осей и побегов. Мелкие ксерофильные листья и цветки расположены по периферии подушки. Между отдельными ветвями скапливаются мелкозем, пыль, мелкие камни. В результате некоторые виды растений-подушек приобретают большую компактность и необычайную плотность: по таким растениям можно ходить, как по твердой почве. Таковы Silene acaulis. Gypsophila aretioides, Androsace helvetica, Acantholimon diapensioides. Издали их трудно отличить от валунов. Менее плотны колючие подушки из родов Eurotia, Saxifraga.
Растения-подушки бывают разных размеров (до 1 м в поперечнике) и разнообразных очертаний: полушаровидные, плоские, вогнутые, иногда довольно причудливых форм (в Австралии и Новой Зеландии их называют «растительными овцами»).
Благодаря компактной структуре растения-подушки успешно противостоят холодным ветрам. Поверхность их нагревается почти так же, как и поверхность почвы, а колебания температуры внутри менее выражены, чем в окружающей среде. Отмечены случаи значительного повышения температуры внутри подушки; например, у наиболее распространенного вида высокогорий Центрального Тянь-Шаня Dryadanthe tetrandra при температуре воздуха 10°С внутри подушки температура доходила до 23°С благодаря аккумуляции тепла в этом своеобразном «парнике». В связи с медленным ростом растения-подушки по долговечности вполне сравнимы с деревьями. Так, на Памире подушка Acantholimon hedini диаметром 3 см имела возраст 10—12 лет, при 10 см — 30—35 лет, а возраст крупных подушек достигал не одной сотни лет.
В пределах общей формы растений-подушек существует экологическое разнообразие: например в горах, окружающих Средиземное море, распространены менее компактные по строению ксерофильные «колючие подушки», которые не встречаются высоко в горах, так как малоустойчивы к холоду, но зато очень устойчивы к засухе. Рыхлое строение подушки здесь оказывается более выгодным для растения, чем компактное, так как в условиях летней засухи и сильной инсоляции снижает опасность перегрева ее поверхности. Температура поверхности средиземноморских подушек обычно ниже температуры воздуха благодаря сильной транспирации, а внутри подушки создается особый микроклимат; например, влажность воздуха держится на уровне 70—80% при влажности наружного воздуха 30%. Таким образом, здесь форма подушки — это приспособление к совсем иному комплексу факторов, отсюда и ее иная «конструкция».
Среди других особенностей роста, помогающих растениям преодолевать действие холода, следует еще упомянуть различные приспособления, направленные на углубление зимующих частей растений в почву. Это развитие контрактильных (сократительных) корней — толстых и мясистых, с сильноразвитой механической тканью. Осенью они высыхают и сильно сокращаются в длину (что хорошо заметно по поперечной морщинистости), при этом возникают силы, втягивающие в почву зимующие почки возобновления, луковицы, корни, корневища.
Контрактильные корни встречаются у многих растений высокогорий, тундр и других холодных местообитаний. Они позволяют, в частности, успешно противостоять морозному выпиранию растений из почвы. В последнем случае они не только втягивают почку возобновления, но и ориентируют ее перпендикулярно поверхности, если растение повалено. Глубина втягивания контрактильными корнями варьирует от сантиметра до нескольких десятков сантиметров в зависимости от особенностей растения и механического состава почвы.
Адаптивное изменение формы как защита от холода — явление, ограниченное в основном холодными районами. Между тем действие холода испытывают и растения более умеренных областей. Гораздо более универсальны физиологические способы защиты. Они направлены прежде всего на снижение точки замерзания клеточного сока, предохранение воды от вымерзания и т. д. Отсюда такие особенности холодостойких растений, как повышение концентрации клеточного сока, главным образом за счет растворимых углеводов. Известно, что при осеннем повышении холодостойкости («закаливании») крахмал превращается в растворимые сахара. Другая черта холодостойких растений — повышение доли коллоидно-связанной воды в общем водном запасе.
При медленном снижении температуры растения могут вынести охлаждение ниже точки замерзания клеточного сока в состоянии переохлаждения (без образования льда). Как показывают эксперименты, уровень точек переохлаждения и замерзания тесно связан с температурными условиями обитания. Однако у растений состояние переохлаждения возможно лишь при небольшом холоде (несколько градусов ниже нуля). Гораздо более действенным этот путь адаптации оказывается у других пойкилотермных организмов— насекомых, у которых роль антифризов играют глицерин, трегалоза и другие защитные вещества (открыто зимующие насекомые могут вынести переохлаждение клеточных соков без замерзания до — 30°С).
Многие растения способны сохранять жизнеспособность и в промерзшем состоянии. Есть виды, замерзающие осенью в фазе цветения и продолжающие цвести после оттаивания весной (мокрица — Stellaria media, маргаритка— Bellis perennis, арктический хрен — Cochlearia fenestrata и др.). Ранневесенние лесные эфемероиды («подснежники») в течение короткой вегетации неоднократно переносят весенние ночные заморозки: цветки и листья промерзают до стекловидно-хрупкого состояния и покрываются инеем, но уже через 2—3 ч после восхода солнца оттаивают и возвращаются в обычное состояние. Хорошо известна способность мхов и лишайников переносить длительное промерзание зимой в состоянии анабиоза. В одном из опытов лишайник Cladonia замораживали при —15°С на 110 недель (более двух лет!). После оттаивания лишайник оказался живым и вполне жизнеспособным, у него возобновились фотосинтез и рост. Очевидно, у лишайников в крайне холодных условиях существования периоды такого анабиоза очень длительны, а рост и активная жизнедеятельность осуществляются лишь в короткие благоприятные периоды (причем не каждый год). Такое частое прерывание активной жизни на долгие сроки, по-видимому, объясняет колоссальный возраст многих лишайников, определенный радиоуглеродным методом (до 1300 лет у Rhizocarpon geographicum и Альпах, до 4500 лет у лишайников в Западной Гренландии).
Анабиоз — «крайняя мера» в борьбе растения с холодом, приводящая к приостановке жизненных процессов и резкому снижению продуктивности. Гораздо большее значение в адаптации растений к холоду имеет возможность сохранения нормальной жизнедеятельности путем снижения температурных оптимумов физиологических процессов и нижних температурных границ, при которых эти процессы возможны. Как видно на примере оптимальных температур для фотосинтеза и его нижних температурных порогов, эти явления хорошо выражены у растений холодных местообитаний. Так, у альпийских и антарктических лишайников для фотосинтеза оптимальна температура около 5°С; заметный фотосинтез удается обнаружить у них даже при —10°С. При сравнительно низких температурах лежит оптимум фотосинтеза у арктических растений, высокогорных видов, ранневесенних эфемероидов. Зимой при отрицательных температурах способны к фотосинтезу многие хвойные древесные породы. У одного и того же вида температурные оптимумы фотосинтеза связаны с изменением условий: так, у альпийских и арктических популяций травянистых многолетников — Охуria digyna, Thalictrum alpinum и других видов они более низкие, чем у равнинных. Показательно в этом отношении и сезонное смещение оптимума по мере повышения температуры от весны к лету и снижения от лета к осени и зиме.
При низких температурах для растений чрезвычайно важно сохранить достаточный уровень дыхания — энергетической основы роста и репарации возможных повреждений холодом. На примере ряда растений памирских высокогорий показано, что в этих условиях довольно интенсивное дыхание сохраняется после действия температуры от —6 до —10°С.
Еще один пример устойчивости физиологических процессов к холоду— зимний и предвесенний подснежный рост у растений тундр, высокогорий и других холодных местообитаний с коротким вегетационным периодом, обусловленным заблаговременной подготовкой. Это явление чрезвычайно ярко выражено у эфемероидов лесостепных дубовых лесов (пролески — Scilla sibirica, хохлатки — Corydalis halleri, гусиного лука — Gagea lutea, чистяка — Ficariaverna и др.), у которых уже в начале зимы начинается рост побегов со сформированными внутри бутонами (вначале в промерзшей почве, а затем над почвой, внутри снежного покрова. Не прекращается у них зимой и формирование генеративных органов. По мере приближения сроков снеготаяния скорость подснежного роста заметно возрастает. В пору раннего «предвесенья», когда лес кажется еще совсем безжизненным, под снеговым покровом над почвой уже возвышаются тысячи ростков пролески и гусиного лука, достигающих к этому времени 2—7 см высоты и готовых начать цветение, как только сойдет снег. Образование хлорофилла у ранневесенних эфемероидов также начинается при низких температурах порядка 0°С, еще под снегом.
Экологические различия холодостойкости растений. В экологии и экологической физиологии в качестве одного из показателей устойчивости к холоду используется способность растения переносить низкую температуру в экспериментальных условиях в течение определенного срока. Накоплено много данных, позволяющих сравнивать растения различных по температурным условиям местообитаний. Однако эти данные не всегда строго сравнимы, поскольку температура, которую способно вынести растение, в числе прочих причин зависит и от продолжительности ее действия (так, небольшой холод порядка —3—5°С умеренно теплолюбивое растение способно вынести в течение нескольких часов, но та же температура может оказаться губительной, если будет действовать несколько суток), В большинстве экспериментальных работ принято охлаждение растений в течение суток или близкого срока.
Как видно из нижеследующих данных и, холодостойкость растений весьма различна и зависит от условий, в которых они обитают.
Один из крайних примеров холодостойкости — так называемый «криопланктон». Это снежные водоросли, живущие в поверхностных слоях снега и льда и при массовом размножении вызывающие его окрашивание («красный снег», «зеленый снег» и т. д.). В активных фазах они развиваются при 0°С (летом на оттаявшей поверхности снега и льда). Пределы устойчивости к низким температурам от —36°С у Chlamydomonas nivalis до —40, —60°С у Pediastrutn boryanum, Hormidium flaccidum. Столь же велика холодостойкость фитопланктона полярных морей, нередко зимующего в корке льда.
Большой холодостойкостью отличаются альпийские карликовые кустарнички — Rhododendron ferrugineum, Erica carnea и др. (—28, —36°С), хвойные древесные породы: так, для сосны Pinus strobus в Тирольских Альпах в экспериментах отмечена рекордная температура: —78°С.
Совсем небольшая холодостойкость у растений тропических и субтропических областей, где они не испытывают действия низких температур (за исключением высокогорий). Так, для водорослей тропических морей (особенно мелководных районов) нижняя температурная граница лежит в пределах 5—14°С (вспомним, что для водорослей арктических морей верхняя граница составляет 16°С). Саженцы тропических древесных пород гибнут при 3—5°С. У многих тропических термофильных растений, например декоративных оранжерейных видов из родов Gloxinia, Coleus, Achimenes и др., понижение температуры до нескольких градусов выше нуля вызывает явления «простуды»: при отсутствии видимых повреждений через некоторое время останавливается рост, опадают листья, растения завядают, а затем и гибнут. Известно это явление и для теплолюбивых культурных растений (огурцов, томатов, фасоли).
Очень невелика устойчивость к холоду у термофильных плесневых грибов из родов Mucor, Thermoascus, Anixia и др. Они гибнут за три дня при температуре 5—6°С и даже температуру 15—17°С не могут выносить дольше 15—20 дней.
В зависимости от степени и специфического характера холодостойкости можно выделить следующие группы растений.
5.1. Нехолодостойкие растения
К этой группе относятся все те растения, которые серьезно повреждаются уже при температурах выше точки замерзания: водоросли теплых морей, некоторые грибы и многие листостебельные растения тропических дождевых лесов.
5.2. Неморозостойкие растения
Эти растения хотя и переносят низкие температуры, но вымерзают, как только в тканях начинает образовываться лед. Неморозостойкие растения защищены от повреждения только средствами, замедляющими замерзание. В более холодное время года у них повышается концентрация осмотически активных веществ в клеточном соке и в протоплазме, а также переохлаждаемость, что предотвращает или замедляет образование льда при температурах примерно до —7°С, а при постоянном переохлаждении и до более низких температур. В период вегетации все листостебельные растения неморозостойки. В течение всего года чувствительны к образованию льда глубоководные водоросли, холодных морей и некоторые пресноводные водоросли, тропические и субтропические древесные растения и различные виды из умеренно-теплых районов.
5.3. Льдоустойчивые» растения
В холодное время года эти растения переносят внеклеточное замерзание воды и связанное с ним обезвоживание. Устойчивыми к образованию льда становятся некоторые пресноводные водоросли и водоросли приливной зоны, наземные водоросли, мхи всех климатических зон (даже тропической) и многолетние наземные растения областей с холодной зимой. Некоторые водоросли, многие лишайники и различные древесные растения способны чрезвычайно сильно закаливаться; тогда они остаются без повреждений и после продолжительных суровых морозов, и их можно охлаждать даже до температуры жидкого азота.
ЗАКЛЮЧЕНИЕ
Удивительная гармония живой природы, ее совершенство создаются самой природой: борьбой за выживание. Формы приспособлений у растений бесконечно разнообразны. Весь растительный мир со времени своего появления совершенствуется по пути целесообразных приспособлений к условиям обитания.
Растения – пойкилотермные организмы. Повреждения начинаются на молекулярном уровне с нарушений функций белков и нуклеиновых кислот. Температура - это фактор, серьезно влияющий на морфологию и физиологию растений, требующий изменений в самом растении, которые могли бы приспособить его. Адаптации растений к разным температурным условиям даже в пределах одного вида различны.
При высоких температурах выявлены такие адаптации, как густое опушение листьев, блестящая поверхность, уменьшение поверхности, поглощающей радиацию, изменение положения по отношению к источнику тепла, усиление транспирации, высокое содержание защитных веществ, сдвиг температурного оптимума активности важнейших ферментов, переход в состояние анабиоза, занятие микрониш, защищенных от инсоляции и перегрева, сдвиг вегетации на сезон с более благоприятными тепловыми условиями.
Адаптации к холоду таковы: опушение почечных чешуй, толстая кутикула, утолщение пробкового слоя, опушение листьев, закрывание розеточных листьев ночью, развитие карликовости, развития стелящихся форм, подушковая форма роста, развитие контрактильных корней, повышение концентрации клеточного сока, повышение доли коллоидно-связанной воды, анабиоз
По различной термоустойчивости выделяются виды: нехолодостойкие, неморозоустойчивые, льдоустойчивые, нежаростойкие, жаровыносливые зукариоты, жаровыносливые прокариоты.
ЛИТЕРАТУРА
Александров В.Я. Клетки, макромолекулы и температура. Л.: Наука, 1975г. 328 с
Вознесенский В. Л., Рейнус Р. М . Температура ассимилирующих органов пустынных растений // Бот. журн., 1977; т. 62. N 6
Горышина Т. К. Ранневесенние эфемероиды лесостепных дубрав. Л., Изд-во Ленингр. ун-та. 1969
Горышина Т.Н. Экология растений уч. Пособие для ВУЗов, Москва, В. школа, 1979г. 63-102с.
Культиасов И.М. Экология растений М.: Изд-во московского ун-та, 1982 33-89с.
Лархер В. Экология растений М.: Мир 1978 г. 283-324c.
Максимов Н. А. Избранные работы по засухоустойчивости и зимостойкости растений М.:Изд-во АН-СССР.-1952 т. 1-2
Полевой В.В. Физиология растений 1978г. 414-424с.
Селянинов Г. Т. К методике сельскохозяйственной климатологии. Труды по с.-х. метеорологии, 1930, т. 22
Тихомиров Б. А. Очерки по биологии растений Арктики. Л., Изд-во АН СССР, 1963
Туманов И. И. Причины гибели растений в холодное время года и меры её предупреждения. М., Знание, 1955
Похожие работы
... которой создается более теплый микроклимат; у обитателей тундры возникают стелящиеся, или шпалерные, и карликовые формы. По отношению к теплу выделяют следующие экологические группы: Эвритермные и стенотермные организмы (см. предыдущий урок) Термофилы и криофилы (теплолюбивые и холодолюбивые) По степени адаптации к условиям дефицита тепла различают нехолодостойкие (гибнут при температуре ...
... , опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, ...
... сумеречного освещения в водных глубинах и пещерах. В разных местообитаниях различаются не только интенсивность света, но и его спектральный состав, продолжительность освещения, пространственное и временное распределение света разной интенсивности и т. д. Соответственно, разнообразны и приспособления растений к жизни при том или ином световом режиме. Экологические группы растений по отношению к ...
... . У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10—100 раз. В пролине ...
0 комментариев