Навигация
1.2.3 Разложение гемицеллюлоз
Гемицеллюлозы – полисахариды, сахара и уроновые кислоты – присутствуют во всех растительных тканях. Ксилан, относящийся к гемицеллюлозам, – полимер ксилозы, занимает по количеству в растениях второе место после целлюлозы. В древесине хвойных его 12%, лиственных деревьев – до 25%.
Разложение гемицеллюлоз – процесс неспецифический и осуществляется многими микроорганизмами. Большое место в этом процессе занимают грибы, в частности фермент ксиланаза есть у многих грибов, как микромицетов, так и высших базидиальных грибов, многие из которых являются типичными сапротрофами на растительном опаде (Мирчинк, 1988).
1.2.4 Разложение пектиновых веществ
Среди грибов имеются активные разлагатели пектина, который также является существенным компонентом растительного опада. Пектин образует в растениях межклеточное вещество, из которого состоят так называемые срединные пластинки, соединяющие между собой отдельные клетки растения. Они придают тканям прочность. Пектин представляет собой высокомолекулярное соединение углеводной природы – полисахарид, в котором метоксилированные остатки галактуроновой кислоты связаны между собой β-1,4-глюкозидными связями.
В растениях пектиновые вещества присутствуют в виде нерастворимого протопектина в соединения с другими полисахаридами клеточной оболочки.
Действие пектиназ проявляется в размягчении ткани и распаде ее на отдельные клетки. Существует несколько типов ферментов-пектиназ в зависимости от специфики их воздействия на субстрат: протопектиназы вызывают превращение нерастворимого протопектина в растворимый протопектин, расщепляют связи между метоксилированной полигалактуроновой кислотой и связанным с ней арабаном и ксиланом; пектинэстеразы гидролизуют метоксильные молекулы растворимого пектина (гидролитическое отщепление метоксильных групп от растворимого пектина), в результате чего образуется полигалактуроновая кислота и метиловый спирт; полигалактуроназы гидролизуют β-1,4-глюкозидные связи пектиновой кислоты до свободных галактуроновых кислот. Встречаются главным образом у различных видов бактерий и грибов.
Многие грибы образуют пектинолитические ферменты. Высокая пектинолитическая активность обнаружена у некоторых эпифитных грибов, главным образом Aureobasidium pullulans и видов Cladosporium. Пектинолитические грибы занимают значительное место среди типичных представителей лесной подстилки – это виды родов Cladosporium, Alternaria, Aposphaeria, Penicillium, фитопатогенные грибы родов Fusarium, Verticillium, Botrytis cinerea, Sclerotinia sclerotiorum.
Ферментативное разрушение пектиновых веществ в растениях имеет значение в патогенезе некоторых заболеваний. Фитопатогенные грибы разрушают пектин срединной пластинкой и пектаты в первичных клеточных оболочках, что приводит к изменению их физико-химических свойств и создает условия для внедрения паразита, а также в результате действия пектинэстеразы образуются вещества – полигалактурониды, способные закупоривать сосуды, что в конечном итоге приводит к увяданию растений.
Существенное значение разрушения пектиновых веществ грибами имеет при разложении растительного опада. Практическое использование пектиназ грибов – применение в пищевой промышленности при приготовлении фруктовых соков для их осветления, а также при мочке льна (Мирчинк, 1988).
1.2.5 Разложение лигнина
Грибы – почти единственные разрушители лигнина. Способность грибов осуществлять глубокое разрушение лигнина представляет собой уникальное явление.
Лигнин – наиболее распространенное в природе полимерное циклическое соединение. В наибольшем количестве лигнин содержится в древесине и древесном опаде. Содержание его в опаде хвойных пород составляет 28 – 34%, лиственный пород – 18 – 28%. В химическом отношении лигнин не является индивидуальным веществом с вполне определенными свойствами и составом.
Исследования, касающиеся микробной деградации лигнина, относятся к одной из наиболее сложных биологических проблем, поскольку лигнин пока не может быть точно определен как химическое вещество. Также не могут быть точно определены и промежуточные реакции его биологического превращения. Наиболее активные группы микроорганизмов, разрушающих лигнин, принадлежат к древоразрушающим базидиомицетам, вызывающим белую гниль. Однако до настоящего времени неизвестны полностью все стадии ферментативных реакций в процессе разложения лигнина, то есть известны далеко не все ферменты, осуществляющие этот процесс.
Отличия деградации лигнина от деградации других полимеров заключается в том, что такие полимеры, как протеины, полисахариды, нуклеиновые кислоты, состоят из регулярно повторяющихся единиц, в то время как лигнин состоит из различных мономеров, имеющих различные типы связей. Большинство микроорганизмов, воздействующих на лигнин, вызывают в нем очень незначительные изменения, которые проявляются в основном в уменьшении числа метоксильных групп и очень слабой потере в весе. Некоторые сумчатые и несовершенные грибы могут расти на средах, содержащих препарат лигнина в качестве единственного источника углерода, такие, как Fusarium lactis, F. nivale и некоторые другие, но они не вызывают существенных изменений в молекуле лигнина. Вещества, представляющие собой производные лигнина, – ванилин, сиреневый альдегид и другие, используются грибами родов Chaetomella, Coniothyrium, Cylindrocarpon, Torula, Hormiscium.
Исследования последних лет показывают, что полное разложение лигнина с разрушением ароматического кольца могут осуществлять только базидиальные древоразрушающие грибы, вызывающие белую гниль: Coriolus versicolor, Fomes fomentarius, и некоторые подстилочные базидиомицеты, такие, как виды родов Collybria, Marasmius, Mycena.
Высокая молекулярная масса и низкая растворимость лигнина препятствуют его прямой ассимиляции микроорганизмами. Предварительно происходит его расщепление экзоферментами во внешней среде. Своеобразное химическое строение лигнина делает его труднодоступным для ферментных систем микроорганизмов. Многие микроорганизмы обладают ферментными системами, способными к превращению простых ароматических соединений. Однако полизамещенные ароматические соединения – мономеры лигнина – доступны лишь немногим представителям отдельных родов, так как для их разложения требуются весьма редкие у микроорганизмов ферменты деметилирования и декарбоксилирования ароматических структур. Необычен для микроорганизмов и характер связей между мономерами, некоторые из них не имеют аналогий среди соединений, типичных для живой клетки. Они совершенно недоступны для ферментов – деполимераз микроорганизмов, легко разрушающих белковые, полисахаридные, полинуклеиновые молекулы.
Способность многих грибов разлагать клетчатку и лигнин определяет их активное участие в разложении растительного опада. В растительном опаде в древесине лигнин и целлюлоза образуют природный комплекс, в котором его составляющие структурно и химически связаны между собой. Разложение лигноцеллюлозного комплекса грибами установлено серией экспериментов по разложению растительного опада, проведенных В.Я. Частухиным. Он разработал специальную методику для лабораторного изучения процессов распада, которая позволила создать модель процессов, происходящих в природе, в условиях точно контролируемых опытов.
По характеру воздействия на опад выделяют три группы подстилочных сапротрофов (Мирчинк, 1988):
I группа – грибы, преимущественно разлагающие клетчатку, – Lepiota procera.
II группа – грибы, преимущественно разлагающие лигнин, – Phallus impudicus.
III группа – грибы, осуществляющие смешанное воздействие, – Collybria, Marasmius, Clitocybe.
Г. Линдбергом были проведены опыты по определению интенсивности разложения целлюлозы и лигнина подстилочными сапротрофами, принадлежащими к различным видам Marasmius, непосредственно в растительном опаде с точным учетом исходного и конечного содержания этих продуктов в субстрате. Почти все виды Marasmius оказались способными разлагать лигнин, но интенсивность разложения лигнина у разных видов различна. Также было различным отношение количеств разложенных лигнина и клетчатки. Таким образом, внутри рода Marasmius можно выделить группы, преимущественно разлагающие лигнин, клетчатку или оба компонента вместе (Мирчинк, 1988).
Похожие работы
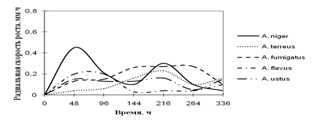
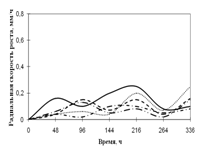
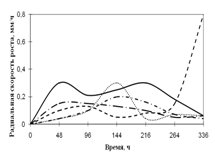
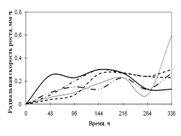
... просмотренных. Наблюдение проводят на 1, 2, 3, 7, 15-е сутки. Стекла микроскопировали, учитывая прорастание спор (%), длину проростков (в мкм) (Кураков, 2001). ГЛАВА 3. ИССЛЕДОВАНИЕ РОСТА МИКРОМИЦЕТОВ НА РАЗЛИЧНЫХ СУБСТРАТАХ Объектами исследования явились 10 штаммов коллекционных микроскоскопических грибов родов Aspergillus: A. niger, A. ustus, A. terreus, A. flavus, A. fumigatus, а также ...
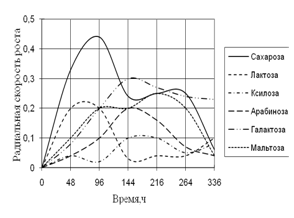
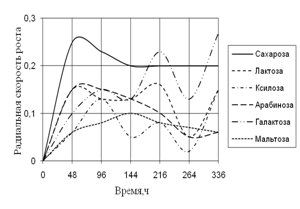
... 5 видов разлагали хитин, 8 – выделяли тирозиназу и лакказу (Parton, 1988). Отмечают высокую целюлозолитическую активность у штаммов видов Trichoderma, Aspergillus niger и Penicillium miszynsky, выделенных из водных местообитаний (Hopper, 1995). Микроскопические грибы могут прямо или косвенно влиять на активность и развитие других гидробионтов. Пути воздействия могут быть различными – от прямого ...
... города Чернигова мною было собрано 13 видов дереворазрушающих грибов. Эти грибы были собраны в 3 – х географических пунктах окрестностей города, а именно Массаны, Яцево, район р. Десна. Распространение трутовиковых грибов в природных условиях определяется рядом факторов, важнейшим из которых служит субстрат, т.е. соответствующая древесина. Способность некоторых видов развиваться только на живых ...
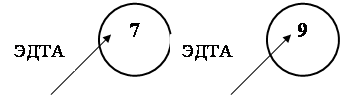
... к ассимиляции ЭДТА и глюкозы в процессе длительного культивирования с добавлением субстрата В предыдущем разделе было показано, что ассимиляция глюкозы бактериальным штаммом LPM-4 индуцируется в процессе деградации ЭДТА, а кометаболизм ЭДТА и глюкозы у штамма LPM-4 не оказывает влияния на деградацию ЭДТА. В данном разделе целью работы было исследование: 1) сохраняется ли способность клеток ...
0 комментариев