Навигация
Использование микромицетами различных природных веществ в качестве единственного источника углерода
56057
знаков
0
таблиц
7
изображений
1.2 Использование микромицетами различных природных веществ в качестве единственного источника углерода
1.2.1 Разложение целлюлозы
Целлюлозолитические системы, состав и активность их отдельных компонентов, продуцируемые различными микроорганизмами, варьируют в широких пределах. В настоящее время наиболее перспективными продуцентами целлюлаз с точки зрения промышленного использования являются грибы следующих родов: Aspergillus, Coriolus, Eupenicillium, Fusarium, Penicillium, Physarium, Sporotrichum, Trichoderma, Verticillium.
Среди грибов, способных расщеплять кристаллическую целлюлозу, только некоторые продуцируют полные внеклеточные целлюлолитические системы (эндо- и экзоглюканазы, β-глюкозидазу). Среди них Trichoderma viride, T. reesei, T. koningii, Penicillium funiculosum, Fusarium solani. Для культуральной жидкости большинства других грибов этой группы характерно отсутствие экзоглюканазы, то есть эти грибы могут деградировать более аморфные формы целлюлозы.
Деградация высокоупорядоченной формы целлюлозы осуществляется благодаря синергическому действию комплекса целлюлолитических ферментов. При любой комбинации экзо- и эндоглюканаз Trichoderma koningii, Fusarium solani, Penicillium и Funiculosum отмечается выраженный синергизм. Однако синергизм между экзоглюканазами этих грибов и эндоглюканазами грибов, не продуцирующих экзоглюканазу (Myrothecium verrucaria), не выявлен. Нет также синергизма между экзоглюканазами грибов и эндоглюканазами рубцовых бактерий. Последнее указывает на существенные различия целлюлолитических систем грибов и бактерий.
В природе в процессе фотосинтеза производится большое количество целлюлозы, в результате возникли многие виды целлюлолитических микроорганизмов. В почве целлюлозная и гемицеллюлозная части биомассы разлагаются интенсивнее, чем лигнин, и быстро метаболизируются почвенными микроорганизмами. Показано, что целлюлаза из Trichoderma viride образует с гуминовыми кислотами комплекс, стабильный в почвенных условиях. При внесении азота в почву разложение лигнина и целлюлозы ускоряется. Добавление глюкозы вызывает обратный эффект. Окончательный продукт деградации целлюлозы – углекислый газ, но если процесс протекает в анаэробной среде, образуется также метан (Марьиновская, 2006).
Содержание целлюлозы в древесном опаде составляет 34 – 59%, несколько меньше содержание гемицеллюлоз и пектина (Мирчинк, 1988).
Целлюлоза является линейным полимером d-глюкозы. Остатки глюкозы в молекуле клетчатки, как и в молекуле целлобиозы связаны β-гликозидной связью. Поэтому клетчатку можно рассматривать как полимер целлобиозы. Norman A.G. и Fuller W.H. (1942) считают, что большинство грибов способно усваивать клетчатку. Несмотря на то, что использование клетчатки грибами имеет большое значение в круговороте веществ в природе, процесс этот изучен далеко не полно.
Campbell W.G. высказал предположение, что первым этапом использования клетчатки грибами является не гидролиз, а окисление. Гидролиз клетчатки можно схематически представить следующим образом клетчатка→целлодекстрины→целлотетроза→целлобиоза→d-глюкоза. Ферменты грибов, расщепляющие целлюлозу еще мало изучены.
Способность грибов усваивать клетчатку колеблется в очень широких пределах. В общем, усвоение клетчатки происходит медленнее, чем усвоение глюкозы. Это обуславливается, по всей вероятности, нерастворимостью клетчатки, в связи с которой действие целлюлазы ограничено поверхностью вещества. Возможно также, что синтез целлюлазы происходит сравнительно медленно (Лилли, 1957).
Основными источниками клетчатки для грибов в природных условиях служат древесина и различные растительные остатки. Хотя основной частью древесины и растительных остатков является именно клетчатка, тем не менее в них, помимо клетчатки, всегда содержатся гемицеллюлоза, камеди, таннины и лигнин. Грибы, разрушающие древесину, обычно подразделяют на виды, вызывающие белую гниль, и виды, вызывающие коричневую гниль. Грибы – возбудители коричневой гнили разрушают предпочтительно клетчатку. Грибы, разрушающие неклетчатковые составные части растительных тканей, вызывают белую гниль. Виды, относящиеся ко второй группе, по-видимому, значительно более многочисленны, чем виды грибов, вызывающие коричневую гниль. По данным Ноблеса М.К. (1948), к грибам, вызывающим белую гниль, относятся следующие виды: Armillaria mellea, Ganoderma lobatum, Lenzites betulinus, Pleurotus ostreatus, Polyporus abietinus, P. cinnabarinus, P. pargamenus. К немногим видам, вызывающим коричневую гниль, принадлежат: Daedalea quercina, Lentinus lepideus, Lenzites trabea, Merulius lacrymans и Trametes americana (Лилли, 1957).
1.2.2 Разложение крахмала
Как и целлюлоза, крахмал является полимером d-глюкозы. Остатки глюкозы в его молекуле соединены между собой α-гликозидной связью, поэтому основной структурной единицей молекулы крахмала, как и молекулы гликогена, следует считать мальтозу. Крахмал состоит из двух различных соединений. Молекулы одного из них, называемого амилазой, имеют неразветвленную углеродную цепочку, тогда как другое соединение, с разветвленной углеродной цепочкой, получило название амилопектина. Зеленые растения синтезируют крахмал, животные и грибы образуют гликоген. Ферментативный гидролиз крахмала может быть схематически представлен следующим образом: крахмал→декстрины→мальтоза→d-глюкоза. Декстрины, имеющие разветвленную углеродную цепочку, лишь частично гидролизуются амилазой. Декстрины с неразветвленной углеродной цепочкой полностью превращаются в мальтозу (Мирбек, 1948).
Крахмал нерастворим в воде. Лишь грибы, образующие амилазу, обладают способностью усваивать крахмал. Существует немало грибов, неспособных развиваться на средах с крахмалом, однако большинство из них может усваивать этот полисахарид. Волконский (1934) установил, что 26 различных изученных им видов и штаммов оомицетов из числа сапролегниевых усваивали как крахмал, так и продукты его гидролиза (декстрины, мальтозу и глюкозу), но не были способны ассимилировать 13 других источников углерода, включая сюда и фруктозу. Позднее Марголин (1942) показал, что 19 из 21 вида грибов, усваивающих мальтозу, обладали также способностью использовать и декстрин (Лилли, 1957).
Похожие работы
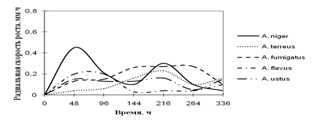
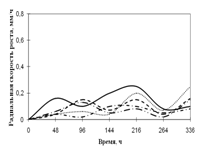
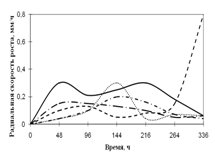
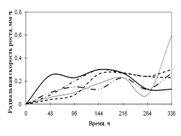
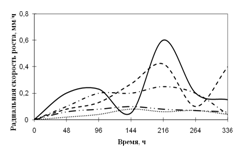
... просмотренных. Наблюдение проводят на 1, 2, 3, 7, 15-е сутки. Стекла микроскопировали, учитывая прорастание спор (%), длину проростков (в мкм) (Кураков, 2001). ГЛАВА 3. ИССЛЕДОВАНИЕ РОСТА МИКРОМИЦЕТОВ НА РАЗЛИЧНЫХ СУБСТРАТАХ Объектами исследования явились 10 штаммов коллекционных микроскоскопических грибов родов Aspergillus: A. niger, A. ustus, A. terreus, A. flavus, A. fumigatus, а также ...
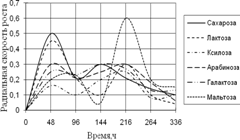
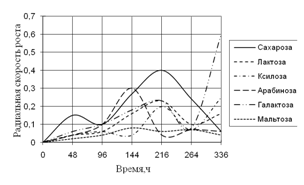
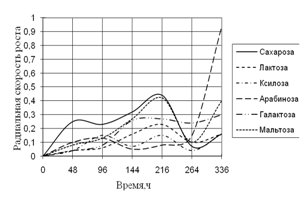
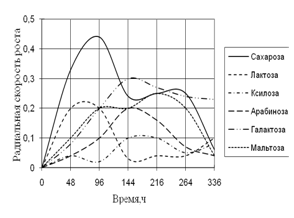
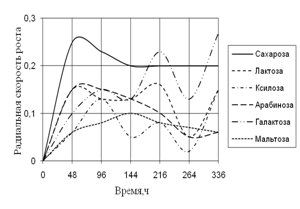
... 5 видов разлагали хитин, 8 – выделяли тирозиназу и лакказу (Parton, 1988). Отмечают высокую целюлозолитическую активность у штаммов видов Trichoderma, Aspergillus niger и Penicillium miszynsky, выделенных из водных местообитаний (Hopper, 1995). Микроскопические грибы могут прямо или косвенно влиять на активность и развитие других гидробионтов. Пути воздействия могут быть различными – от прямого ...
... города Чернигова мною было собрано 13 видов дереворазрушающих грибов. Эти грибы были собраны в 3 – х географических пунктах окрестностей города, а именно Массаны, Яцево, район р. Десна. Распространение трутовиковых грибов в природных условиях определяется рядом факторов, важнейшим из которых служит субстрат, т.е. соответствующая древесина. Способность некоторых видов развиваться только на живых ...
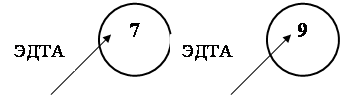
... к ассимиляции ЭДТА и глюкозы в процессе длительного культивирования с добавлением субстрата В предыдущем разделе было показано, что ассимиляция глюкозы бактериальным штаммом LPM-4 индуцируется в процессе деградации ЭДТА, а кометаболизм ЭДТА и глюкозы у штамма LPM-4 не оказывает влияния на деградацию ЭДТА. В данном разделе целью работы было исследование: 1) сохраняется ли способность клеток ...
0 комментариев