Экология биологического круговорота
Химический состав живого вещества как следствие избирательного перемещения веществ в биологическом круговороте
Незамкнутость биологического круговорота
Звенья преобразования веществ в биологическом круговороте
Классификации круговоротов
Биологический круговорот в таежных сообществах
Хозяйственная деятельность и особенности биогеохимического цикла углерода в лесных экосистемах
Навигация
Экология биологического круговорота
Лесной тип биологического круговорота
75424
знака
4
таблицы
0
изображений
1. Экология биологического круговорота
1.1 Энергетическое обеспечение биологического круговорота и трофические цепи/сети
Все преобразования веществ в процессе круговорота требуют затрат энергии. Ни один живой организм не продуцирует энергию – она может быть получена только извне. В современной биосфере важнейшим источником энергии, утилизируемой в биологическом круговороте, является энергия солнечного излучения. Соответственно первый этап использования и преобразования энергии – фотосинтез, в процессе которого создаются вещества для построения тела растительного организма. Энергия, полученная в виде ФАР, в процессе фотосинтеза преобразуется в энергию химических связей. Процесс аккумуляции энергии в организме фотосинтетиков сопряжен с увеличением массы организма, этот прирост массы обозначается как первичная продукция.
Поскольку не вся полученная фотосинтетиками энергия накапливается в виде первичной продукции, часть её рассеивается в форме тепла. В свою очередь, часть энергии, накопленной в биомассе, расходуется на процессы жизнедеятельности, что ведёт к уменьшению биомассы (потери на дыхание). В результате в виде накопленной биомассы (чистая первичная продукция) аккумулируется лишь относительно небольшая часть полученной организмом продуцента солнечной энергии.
Если энергию солнечного излучения принять за 100%, то лишь 15% ее достигает поверхности Земли и только 1% связывается в виде органического вещества растительности. Из суммы связанной в процессе продукции энергии около половины расходуется на жизненные процессы (потери на дыхание). Оставшиеся 50% аккумулированной энергии составляет рост биомассы. Таким образом, чистая продукция соответствует примерно 0,5% солнечной энергии, падающей на Землю (Шилов, 2000).
Часть энергии накопленной в результате фотосинтеза растений используется в качестве пищи организмами-гетеротрофами. По приблизительным расчетам, в пищу фитофагами изымается около 40% фитомассы, оставшиеся 60% означают реальную массу растительности в экосистеме.
Усвоенная энергия, за вычетом энергии, содержащейся в выведенных из организма секретах, составляет метаболированную энергию. Часть её выделяется в виде тепла в процессе переваривания пищи и либо рассеивается, либо используется на терморегуляцию. Оставшаяся энергия подразделяется на энергию существования, которая расходуется на различные формы жизнедеятельности, и продуктивную энергию, аккумулирующуюся в виде массы нарастающих тканей, энергетических резервов, половых продуктов. Энергия, накопленная в тканях тела гетеротрофа, составляет вторичную продукцию экосистемы, которая может быть использована в пищу консументами высших порядков.
Подобным образом энергия расходуется на всех гетеротрофных этапах круговорота, т. е. в организмах, последовательно использующих в пищу биомассу предыдущих трофических уровней. В результате количество энергии, доступной для потребления, прогрессивно падает по ходу повышения трофических уровней, что лежит в основе относительно небольшой длины пищевых цепей.
На фоне биологического круговорота веществ потоки энергии однонаправлены: первично аккумулированная в тканях продуцентов энергия постепенно рассеивается в виде тепла на всех этапах трофических цепей. Однако на всех этапах идет и синтез вещества, а вместе с тем аккумуляция энергии в химических связях (Шилов, 2000).
Именно благодаря однонаправленному движению энергии по трофическим цепям осуществляется движение атомов, организуются экосистемы. Это отражено в законе биологического круговорота элементов в ландшафте Вернадского-Полынова (Перельман, 1975): химические элементы в ландшафте совершают круговороты, в ходе которых многократно поступают в живые организмы («организуются») и выходят из них («минерализуются»). При этом происходит поглощение и выделение энергии, совершается химическая работа, изменяется степень разнообразия. Поступательное развитие ландшафтов (экосистем) осуществляется через систему круговоротов.
Благодаря энергии, поступающей в экосистему, происходит дифференциация живого вещества на несколько трофических уровней, каждый последующий уровень существует за счет части энергии предыдущего. Формируется трофическая структура экосистемы.
Все многообразие видов, входящих в состав экосистем, с необходимостью разделяется на три принципиальных экологических группы – продуцентов, консументов, редуцентов. В конкретных биоценозах они представлены популяциями многих видов, состав которых специфичен для каждого конкретного сообщества. Равнозначные виды образуют определенный трофический уровень, а взаимоотношения между видами – систему цепей питания (трофическая сеть).
Группа видов продуцентов образует уровень первичной продукции, на котором утилизируется внешняя энергия и создается масса органического вещества. Первичные продуценты – основа трофической структуры и всего существования всего биоценоза. Биомасса органического вещества синтезированного продуцентами – первичная продукция, а скорость её образования – биологическая продуктивность. Общая сумма биомассы рассматривается в этом случае как валовая продукция, а та её часть, которая определяет прирост, - как чистая продукция. Разница между валовой и чистой продукцией определяется затратами на дыхание.
Консументы I порядка. Этот трофический уровень составлен непосредственными потребителями первичной продукции. В наиболее типичных случаях, когда биомасса продуцентов создается фотоавтотрофами, это фитофаги. Консументы частично используют пищу для обеспечения жизненных процессов, а частично строят на ее основе собственное тело, осуществляя таким образом первый, принципиальный этап трансформации органического вещества, синтезированного продуцентами. Процесс создания и накопления биомассы на уровне консументов обозначается как вторичная продукция.
Консументы II и III порядков. Эти уровни объединяют животных с плотоядным типом питания (зоофаги). В первую группу входят все хищники, поскольку их специфические черты практически не зависят от того, является жертва фитофагом, или плотоядна. Во вторую группу (третий порядок) входят паразиты и «сверхпаразиты», хозяева которых паразиты.
Деление биоценоза на трофические уровни представляет собой лишь общую схему. Действительные формы взаимоотношений сложнее. Существует много видов со смешанным питанием, которые могут относиться к различным трофическим группам.
Переход биомассы с нижележащего трофического уровня на вышележащий связан с потерями вещества и энергии. В среднем считается, что лишь порядка 10% биомассы и связанной в ней энергии переходит с каждого уровня на следующий. В силу этого суммарная биомасса, продукция и энергия уменьшаются по мере восхождения по трофическим уровням. Эта закономерность сформулирована Ч. Элтоном в виде правила экологических пирамид и существует как главный ограничитель длинны трофических цепей (Шилов, 2000).
Прямые пищевые связи типа «растение-фитофаг-хищник-паразит» объединяют в цепи питания, или трофические цепи.
Каждый трофический уровень составлен не одним, а многими конкретными видами. Благодаря видоспецифичности питания увеличение числа видов в биоценозе определяет более полное использование ресурсов на каждом трофическом уровне. Это обстоятельство прямо связано с повышением полноты биогенного круговорота веществ.
Увеличение видового разнообразия выступает как «гарантийный механизм», обеспечивающий надежность круговорота веществ как главной функции экосистем. Суть механизма заключается в том, что монофагия встречается в природе довольно редко; немногочисленны и олигофаги. Большинство животных использует в пищу более или менее широкий набор кормовых объектов. В результате помимо прямых «вертикальных» пищевых связей возникают боковые, объединяющие потоки вещества и энергии двух и более пищевых цепей. Таким путем формируются пищевые трофические сети, в которых множественность цепей питания выступает как приспособление к устойчивому существованию экосистемы в целом: «дублирование» потоков вещества и энергии по большому числу параллельных трофических цепей поддерживает непрерывность круговорота при всегда вероятных нарушениях отдельных звеньев пищевых цепочек.
Рассмотренные выше процессы связаны с синтезом и трансформацией органического вещества в трофических сетях и характеризуют собой так называемые цепи выедания или «пастбищные цепи». Процессы поэтапной деструкции и минерализации органических веществ обычно выводятся в отдельный блок трофической структуры, называемой цепями разложения (детритные цепи).
Вычленение детритных цепей связано, прежде всего, с тем, что минерализация органики практически идет на всех трофических уровнях: и растения и животные в процессе метаболизма редуцируют органическое вещество. Детритные же цепи начинаются с разложения мертвой органики сапрофагами, которые механически, а отчасти и химически подготавливают органическое вещество к действию редуцентов. В наземных экосистемах этот процесс преимущественно сосредоточен в подстилке и почве.
На уровне консументов происходит разделение потока органического вещества по разным группам потребителей: живое органическое вещество следует по цепям выедания, а мертвое – по цепям разложения. В наземных биоценозах цепи разложения имеют очень большое значение в процессе биологического круговорота; в них перерабатывается до 90% прироста биомассы растений, попадающей в эти цепи в виде опада (Шилов, 2000).
Похожие работы
... опять круговорот она может лишь в результате геологических процессов или путем извлечения живым веществом. Однако извлечь нужные живым организмам вещества из земной коры гораздо сложнее, чем из атмосферы. Интенсивность биологического круговорота в первую очередь определяется температурой окружающей среды и количеством воды. Так, например, биологический круговорот интенсивнее протекает во ...
... , являющихся защитными и выполняющих водоохранные, почвозащитные и другие полезные функции. В первую очередь резерваты были отведены на территории Катон-Карагайского государственного национального природного парка ЛГР №12-17. Ведение хозяйства в лесных генетических резерватах осуществляется в соответствии с Типовыми положениями, Лесным кодексом Республики Казахстан и другими нормативными актами. ...
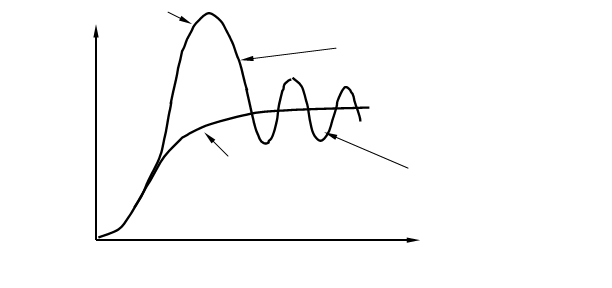

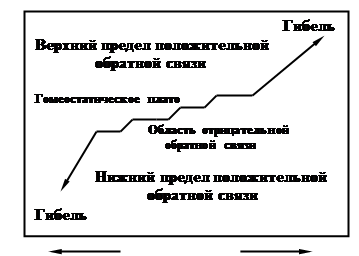
... будет описываться гомеостатическим плато (рис. 6) – областью отрицательных связей, а при нарушении системы начинают преобладать обратные положительные связи, что может привести к гибели системы. Применительно к лесному фитоценозу гомеостаз характеризуется относительно постоянным числом эдификаторов и упрочением связей между всеми ярусами и видами, входящими в биогеоценоз, вселение «чужаков» в ...
... этой подстилки и условиях ее перегнивания. Положительная ролькак источника непосредственного питания и как мертвого покрова, защищающего почву от испарения влаги. 3. Функции и роль подстилки в экосистеме 3.1 Экологическая роль лесных подстилок в миграции техногенных загрязнителей Исследования миграции радиоактивных веществ показали, что после их поступления на поверхность лесной подстилки ...
0 комментариев