Экология биологического круговорота
Химический состав живого вещества как следствие избирательного перемещения веществ в биологическом круговороте
Незамкнутость биологического круговорота
Звенья преобразования веществ в биологическом круговороте
Классификации круговоротов
Биологический круговорот в таежных сообществах
Хозяйственная деятельность и особенности биогеохимического цикла углерода в лесных экосистемах
Навигация
Хозяйственная деятельность и особенности биогеохимического цикла углерода в лесных экосистемах
Лесной тип биологического круговорота
75424
знака
4
таблицы
0
изображений
3.3. Хозяйственная деятельность и особенности биогеохимического цикла углерода в лесных экосистемах
Основными пулами углерода в лесных экосистемах являются: дебрис, пул углерода подстилки, пул углерода лабильного гумуса и пул углерода стабильного гумуса.
Пул дебриса пополняется из пула живой фитомассы за счет отпада. Процесс пополнения пула подстилки из пула живой фитомассы называется опадом. Опад включает сезонный листопад лиственных пород и лиственницы, отмирание хвои у вечнозеленых хвойных, сезонное отмирание травянистых растений напочвенного покрова, отмирание тонких ветвей и поверхностных частей коры, опад генеративных органов деревьев. Кроме того, пул подстилки пополняется и из пула дебриса за счет опада листвы или хвои, тонких ветвей и мелких частей коры усохших деревьев. Как пул дебриса, так и пул подстилки подвергаются гетеротрофной деструкции, в результате чего часть углерода переходит непосредственно в атмосферу. Оставшаяся часть углерода дебриса и подстилки трансформируется в углерод лабильного гумуса почвы. Часть лабильного гумуса минерализуется, в результате чего углерод выделяется в атмосферу, а часть переходит в стабильный гумус. В свою очередь стабильный гумус также подвергается процессам минерализации, а углерод освобождается в атмосферу. В устойчивых экосистемах климатического климакса все перечисленные пулы стремятся к стабилизации, а потоки - к равновесию. Однако далеко не все российские леса могут рассматриваться как равновесные. В первую очередь это относится к лесам Европейской части России, которая за прошедшее столетие испытала многочисленные изменения в землепользовании. По сравнению с Азиатской частью РФ в этом регионе намного выше доля молодых и средневозрастных насаждений, образовавшихся как в результате интенсивной лесоэксплуатации 50-70-х годов, так и в результате зарастания сельскохозяйственных земель (коллективизация 30-х годов, период Великой Отечественной войны, экономический кризис 90-х годов). Существенные изменения детритного компонента углеродного цикла лесов могут произойти при усилении проявлений глобальных изменений климата. При наличии устойчивого потепления произойдут изменения границ климатических климаксов, в результате чего многие ныне устойчивые лесные экосистемы превратятся в стадии сукцессии. Их переход в новое устойчивое состояние, очевидно, займет значительное время, а результатом может стать как потеря углерода, так и накопление его, в первую очередь в пулах лабильного и стабильного гумуса (Кудеяров, Заварзин и др., 2007).
Как показывает моделирование накопления углерода в процессе восстановления климаксовых лесов и при различных хозяйственных нагрузках (различные системы рубок) только для лесов естественного развития рассчитанное соотношение потоков (баланс) углерода положительно и может составить 125-130 т С/га за 200-летний период. При любом из вариантов хозяйственного использования изъятие углерода с древесиной в рассматриваемых авторами лесах не компенсируется созданием новой биомассы; наибольшие потери характерны для практики сплошных рубок (Комаров, Припутина, 2006; Кудеяров, Заварзин и др., 2007).
В настоящее время в Европейской части России отмечается достоверное увеличение продуктивности спелых и перестойных лесов в большинстве таежных регионов. Но, несмотря на то, что факт увеличения приростов демонстрируется довольно часто, основные причины ещё неясны. Комаров, Припутина с соавторами (2006) рассматривают роль увеличения техногенных выпадений азота из атмосферы в несколько раз, как фактор возрастания продуктивности. Моделирование воздействия дополнительных количеств азота при различных сценариях ведения лесного хозяйства привело к следующим выводам:
А) реакции почвы и растительности на уровень азотных выпадений из атмосферы проходят с различной скоростью, почва откликается на изменения быстрее, однако ее буферных способностей оказывается достаточно для того, чтобы растительность ещё достаточно долго поддерживала уровень своей продуктивности;
Б) существует некоторый уровень поступления азота в лесную экосистему, при котором продуктивность экосистемы превышает потери, связанные с лесозаготовками, производимыми по экологически обоснованным критериям, в том числе и по нормативам рубок;
В) даже относительно высокий уровень выпадений азота (20 кг N/га в год) при «хищнических» лесозаготовках уменьшает устойчивость лесной экосистемы и приводит к потере углерода и азота в лесной растительности и почве.
Заключение
Таким образом, почва как многокомпонентная система всегда является центром биологического круговорота. Она обеспечивает его протекание работой различных механизмов. Среди них не только механизм выветривания, снабжающий питательными элементами живые организмы на ранних стадиях круговорота, но и механизм удержания биофильных элементов, что обусловливается глинными минералами. В почве создаются и удерживаются новые биогенные тела природы, причем возобновляемые, такие как подстилка и гумус, чью роль в круговороте трудно переоценить. Последние обеспечивают и причем чрезвычайно длительный период нормальное функционирование наземных экосистем и высокую продуктивность. Почва одновременно может являться транзитной системой для поступления биофильных элементов, участвующих в круговороте. Таким примером является внеландшафтное поступление веществ из атмосферы, а также из почвенно-грунтовых вод или поступление элементов с латеральным потоком. В жестких экологических условиях, например, в северной тайге или тундре не случайно более высокая продуктивность обнаруживается в условиях склоновых, часто полугидроморфных ландшафтов, где осуществляется дополнительное поступление биофильных элементов с латеральным потоком. Поэтому биологический круговорот осуществляется не только на фоне различных петрографических матриц, но и различных условий функционирования почвы. Функционирование почвы может быть не только дополнительным фактором обеспечивающим цикличность круговорота, но и зачастую ведущим, причем на разных уровнях организации биосферы (Васильевская, Богатырев, 2003).
Перспективы развития учения о биологическом круговороте лежат в различных плоскостях, но составляют единое целое. В настоящий период теория несколько опережает экспериментальные и эмпирические наблюдения. Далеко не все идеи, положения и имеющиеся в литературе показатели реализуются в выполняемых в этой области научных исследованиях (Богатырев, 2005).
В заключение приведем несколько фундаментальных показателей по Богатыреву (2005), которые в силу разных обстоятельств ещё не получили должного развития в учении о биологическом круговороте.
В первую очередь – это величина, характеризующая суммарное содержание зольных элементов – речь идет, прежде всего, об истинной, или чистой золе.
Второй важнейший пробел заключен в недостаточном использовании энергетических величин. Довольно малое число исследователей редко пользуется даже косвенным опытом расчетов калориметрических величин, между тем как определение содержания углеводов, белков и жиров позволяет довольно точно рассчитать величины теплотворной способности.
Третий, существенный пробел заключается по-прежнему в отсутствии полной и достоверной информации о продуктивности наземных экосистем. Отметим, что представление о циклах круговорота основывается на однократном определении величин продуктивности и гораздо реже на исследовании динамики процессов продуктивности. Почти уникальными материалами можно считать сведения о круговороте, при котором использовался метод интенсивностей потоков. Между тем, как использование последнего метода изменяет соотношение ведущих элементах, участвующих в круговороте.
Четвертый пробел обусловлен различной степенью исследования наземных экосистем. В целом наименьшей информативностью характеризуются тундровые экосистемы и районы с распространением многолетнемерзлых пород
Пятый пробел обусловлен слабой изученностью биохимического состава структурных составляющих фитомассы, включая подстилку, что в свою очередь не позволяет достаточно достоверно судить, прежде всего, о биогеохимии углерода.
Литература
Алексеенко В. А. Экологическая геохимия. М.: Логос, 2000, 627 с.
Базилевич Н. И. Географические закономерности структуры и функционирования экосистем. – М.: Наука, 1986. – 296 с.
Белянина Л. А. Состав почвенных растворов, почвенно-грунтовых и поверхностных вод территории Центрально-лесного государственного природного биосферного заповедника: Автореф. дис. …канд. биол. наук. – М., 2007. – 25 с.
Богатырев Л. Г. О теоретическом и критериальном информационном обеспечении исследований в области биологического круговорота // Экспериментальная информация в почвоведении: теории, методы получения и пути стандартизации. Москва, 2005.
Васильевская В. Д., Богатырев Л. Г. Функции почв как основного звена в цикле биологического круговорота веществ и устойчивость наземных экосистем. – Структурно-функциональная роль почв и почвенной биоты в биосфере. – М.: Наука, 2003. – С. 174-188.
Гришина Л. А. Биологический круговорот и его роль в почвообразовании. – М.: Изд-во МГУ, 1974. – 127 с.
Добровольский В. В. Основы биогеохимии. – М.: Высш. шк., 1998. – 413 с.
Злотин Р. И., Ходашова К. С. Роль животных в биологическом круговороте лесостепных экосистем. – М.: Наука, 1974. – 200 с.
Казимиров Н. И., Волков А. Д., Зябченко С. С., Иванчиков А. А., Морозова Р. М. Обмен веществ и энергии в сосновых лесах Европейского Севера. – Л.: Наука, 1977. – 304 с.
Казимиров Н. И., Морозова Р. М., Куликова В. К. Органическая масса и потоки веществ в березняках средней тайги. – Л.: Наука, 1978. – 216 с.
Карпачевский Л. О. Экологическое почвоведение. – М.: ГЕОС, 2005 – 335 с.
Касимов Н. С. Базовые концепции и принципы геохимии ландшафтов // Геохимия биосферы. – Смоленск: Ойкумена, 2006. – С. 21-25.
Керженцев А. С. Функциональная экология. – М.: Наука, 2006. – 259 с.
Ковда В. А. Основы учения о почвах. М.: Наука, 1973. Кн. 1 – 447 с., кн 2 – 468 с.
Коломыц Э. Г. Региональная модель глобальных изменений природной среды. – М.: Наука, 2003. – 371 с.
Комаров А. С., Припутина И. В., Михайлов А. В., Чертов О. Г. Биогеохимический цикл углерода в лесных экосистемах России и его техногенные изменения // Почвенные процессы и пространственно-временная организация почв – М.: Наука, 2006. – 568 с.
Перельман А. И. Геохимия ландшафта. – М.: Высш. шк., 1975. – 342 с.
Перельман А. И., Касимов Н. С. Геохимия ландшафта. М.: Астрея, 1999, 768 с.
Кудеяров В. Н., Заварзин Г. А., Благодатский С. А., Борисов А. В. и др. Пулы и потоки углерода в наземных экосистемах России. – М.: Наука, 2007. – 315 с.
Ремезов Н. П., Быкова Л. Н., Смирнова К. М. Потребление и круговорот азота и зольных элементов в лесах Европейской части СССР – М.: Изд-во Моск. Ун-та, 1959. – 282 с.
Родин Л.Е., Ремезов Н.П., Базилевич Н.И. Методические указания к изучению динамики и биологического круговорота в фитоценозах - Л.: Наука, 1968.- 143 с.
Смольянинов И. И. Биологический круговорот веществ и повышение продуктивности лесов. – М.: Лесная промышленность, 1969. – 191 с.
Титлянова А. А., Тесаржова М. Режимы биологического круговорота. – Новосибирск: Наука, 1991. – 150 с.
Трофимов С. Я. Функционирование почв: определение, категории процессов, подходы к типологии // Регуляторная роль почв в функционировании таежных экосистем. – М.: Наука, 2002. – 364 с.
Фокин А. Д. Роль растений в формировании транспортных потоков вещества в почвах // Организация почвенных систем. – Пущино: ИФХБПП РАН, 2007. Т. 1 – С. 50-52.
Чижикова Н. П., Верховец И. А., Лебедева-(Верба), Владыченский А. С. Формирование почв на покровных суглинках в южнотаежной зоне под разными фитоценозами // Почвообразовательные процессы. – М.: Почв. ин-т им В. В. Докучаева, 2006. – 510 с.
Шилов И. А. Экология. – М.: Высш. шк., 2000. – 512 с.
Шишов Л. Л., Кауричев И. С., Большаков В. А. и др. Лизиметры в почвенных исследованиях. – М.: Почв. ин-т им В. В. Докучаева, 1998. – 264 с.
Яшин И. М., Шишов Л. Л., Раскатов В. А. Методология и опыт изучения миграции веществ. М.: Изд-во МСХА, 2001. – 173 с.
Похожие работы
... опять круговорот она может лишь в результате геологических процессов или путем извлечения живым веществом. Однако извлечь нужные живым организмам вещества из земной коры гораздо сложнее, чем из атмосферы. Интенсивность биологического круговорота в первую очередь определяется температурой окружающей среды и количеством воды. Так, например, биологический круговорот интенсивнее протекает во ...
... , являющихся защитными и выполняющих водоохранные, почвозащитные и другие полезные функции. В первую очередь резерваты были отведены на территории Катон-Карагайского государственного национального природного парка ЛГР №12-17. Ведение хозяйства в лесных генетических резерватах осуществляется в соответствии с Типовыми положениями, Лесным кодексом Республики Казахстан и другими нормативными актами. ...

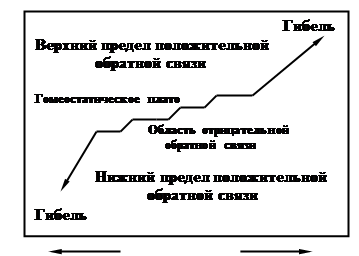
... будет описываться гомеостатическим плато (рис. 6) – областью отрицательных связей, а при нарушении системы начинают преобладать обратные положительные связи, что может привести к гибели системы. Применительно к лесному фитоценозу гомеостаз характеризуется относительно постоянным числом эдификаторов и упрочением связей между всеми ярусами и видами, входящими в биогеоценоз, вселение «чужаков» в ...
... этой подстилки и условиях ее перегнивания. Положительная ролькак источника непосредственного питания и как мертвого покрова, защищающего почву от испарения влаги. 3. Функции и роль подстилки в экосистеме 3.1 Экологическая роль лесных подстилок в миграции техногенных загрязнителей Исследования миграции радиоактивных веществ показали, что после их поступления на поверхность лесной подстилки ...
0 комментариев