Экология биологического круговорота
Химический состав живого вещества как следствие избирательного перемещения веществ в биологическом круговороте
Незамкнутость биологического круговорота
Звенья преобразования веществ в биологическом круговороте
Классификации круговоротов
Биологический круговорот в таежных сообществах
Хозяйственная деятельность и особенности биогеохимического цикла углерода в лесных экосистемах
Навигация
Классификации круговоротов
Лесной тип биологического круговорота
75424
знака
4
таблицы
0
изображений
2.2. Классификации круговоротов
Заключительным этапом при изучении биологического круговорота (БИК) является классификация, которая сводится к упорядочению полученного материала, определению специфики происходящих в БИК процессов и последующему установлению характерных особенностей циклов внутри одной почвенно-биоклиматической зоны или сопоставлению экосистем в зональном аспекте.
Все разнообразные классификации учитывают положение изученного БГЦ в системе почвенно-биоклиматических зон. Вторая особенность классификаций состоит в том, что критерии для классификации выбираются на эмпирической основе. Несомненно, что большинство из них важные, но являются ли они существенными, т.е. достаточными, чтобы отличать БГЦ друг от друга, не всегда ясно. Некоторые показатели БИКа для различных географических зон перекрываются (Васильевская, Богатырев, 2003).
Наиболее широко используемой группой являются данные по продуктивности. Придавая исключительную роль показателям продуктивности, Базилевич (1986) использовала их для оценки структуры и функции наземных экосистем (табл. 26).
Из-за зависимости этих показателей от гидротермического режима, группировка зональных экосистем в таксоны высокого ранга осуществляется в пределах биоклиматических областей одного термического пояса. Для характеристики структуры семейств экосистем используются два критерия: запасы живой фитомассы и запасы мертвой массы, без учета гумуса и торфа.
Основные закономерности структуры экосистем и функционирования сводятся к следующим положениям:
1) изменение величины годичной продукции экосистем по зонам описывается двухвершинной кривой; 2) для интразональных переувлажненных экосистем с севера на юг наблюдается непрерывное возрастание величины годичной продукции; 3) отношение мортмассы к годичной продукции подчинено строгой закономерности, включая зональные и интразональные экосистемы; 4) наибольшие скорости оборота характеризуют экосистемы суббореального и субтропического поясов, что связано, прежде всего, с тепловым балансом земной поверхности.
Показатели продуктивности положены в основу классификации ландшафтов. Так Перельман и Касимов (1999) используют два критерия: биомассу и продуктивность. Таксономия классификации включает группы, типы, семейства. Группы ландшафтов определяются по принадлежности к зоне (тундровая, лесная, степная и т.д.). В ряде случаев выделение групп обусловлено другими факторами, включая позональное их размещение (например, верховые болота в таежной зоне, соровые солончаки в пустынной и т.д.).
В основу выделения типа биогенного ландшафта положен коэффициент К, выводимый из параболической зависимости П = БК, где П - ежегодная продукция, Б - общая биомасса. Для таежной зоны К = 0,54-0,55; для широколиственных лесов - 0,58-0,60; для влажных степей - 0,81. Коэффициент К постоянен при различных величинах П и Б. Тип ландшафта в целом соответствует типам растительного покрова. Внутри ландшафтов, характеризующихся различными биомассой и продукцией, но близким соотношением между ними, выраженным коэффициентом к (П/Б), выделяют три семейства. Например, в пределах таежной зоны выделяются северное, среднее и южное (Васильевская, Богатырев, 2003).
В детальной и получившей наибольшую известность классификации круговорота элементов Н.И. Базилевич используется шесть признаков: 1) принадлежность БГЦ той или иной зональной единице; 2) показатели структуры фитомассы: а) биомасса растений, б) годичный прирост, в) спад, г) истинный прирост, д) подстилка; 3) показатели интенсивности круговорота, устанавливаемые по отношению величины запаса подстилки к величине опада; 4) характер сочетаний химических элементов, потребляемых на построение годичного прироста или возвращаемых с годичным приростом. Типизацию химизма обменных процессов производят по двум ведущим элементам с указанием сопутствующих элементов. Группы типов химизма обменных процессов объединяют в классы по одному преобладающему элементу; 5) величина средней зольности прироста - спада оценивается по средневзвешенному содержанию зольных элементов в 100 г прироста - спада (в %); 6) показатели емкости БИКа оцениваются по величине ежегодно потребляемых на построение прироста (возвращаемых с спадом) зольных элементов и азота (в кг/га). В целях унификации принята десятибалльная шкала числовых показателей.
3 Лесной тип биологического круговорота
3.1. Различия степных и лесных экосистем
Энергетические затраты на первичную продукцию и в целом на биогеоценотические процессы возрастают от среднетаежных растительных формаций к подтайге и широколиственным лесам, а затем снова уменьшаются по направлению к южной лесостепи и северной степи. Максимальное количество энергии используется в хвойно-широколиственных и неморальных лесах, а также в южнотаежных пихтово-ельниках и сосняках.
Аналогичная картина свойственна энергетике групп почв. Практически вся плеяда дерново-подзолистых и серых лесных почв отличаются наибольшими суммарными затратами энергии на почвообразование, между тем как южные черноземы и темно-каштановые почвы развиваются в условиях минимальных затрат.
Стационарные ландшафтно-геофизические эксперименты позволяют выявить причинные механизмы различий между лесными и лугово-степными экосистемами. Так, в одних и тех же зональных условиях типичной лесостепи коэффициент эффективности использования (поглощения) фотосинтетически активной радиации в снытьевой дубраве в два с лишним раза выше, чем в некосимой степи. Кроме того, в лесу имеет место четырехкратное повышение транспирационного коэффициента фитобиоты. Очевидно, более сложная фитоценотическая структура, а также более интенсивная транспирация требуют и больших энергетических затрат. Однако при относительно низком уровне поглощения солнечной энергии травянисто-степная экосистема использует ее более эффективно благодаря тому, что у нее практически выпадает целое звено биологического круговорота – создание скелетной (древесной) многолетней фитомассы, изымающие из годового цикла значительную часть чистой продукции. В результате малый биологический круговорот становится проще, интенсивнее и более замкнут. Образно говоря, с точки зрения устойчивости метаболизма в одних и тех же зональных условиях, допускающих одновременное существование степной и лесной растительности, степь оказывается энергетически более выгодной экосистемой, чем лес (Коломыц, 2003).
Похожие работы
... опять круговорот она может лишь в результате геологических процессов или путем извлечения живым веществом. Однако извлечь нужные живым организмам вещества из земной коры гораздо сложнее, чем из атмосферы. Интенсивность биологического круговорота в первую очередь определяется температурой окружающей среды и количеством воды. Так, например, биологический круговорот интенсивнее протекает во ...
... , являющихся защитными и выполняющих водоохранные, почвозащитные и другие полезные функции. В первую очередь резерваты были отведены на территории Катон-Карагайского государственного национального природного парка ЛГР №12-17. Ведение хозяйства в лесных генетических резерватах осуществляется в соответствии с Типовыми положениями, Лесным кодексом Республики Казахстан и другими нормативными актами. ...
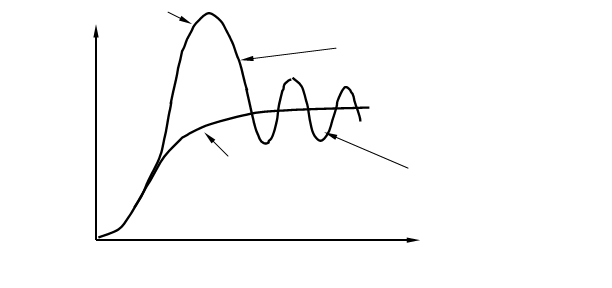

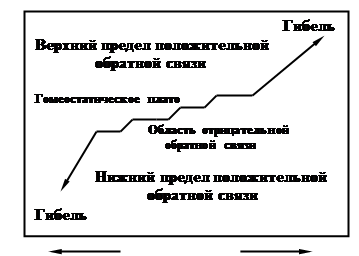
... будет описываться гомеостатическим плато (рис. 6) – областью отрицательных связей, а при нарушении системы начинают преобладать обратные положительные связи, что может привести к гибели системы. Применительно к лесному фитоценозу гомеостаз характеризуется относительно постоянным числом эдификаторов и упрочением связей между всеми ярусами и видами, входящими в биогеоценоз, вселение «чужаков» в ...
... этой подстилки и условиях ее перегнивания. Положительная ролькак источника непосредственного питания и как мертвого покрова, защищающего почву от испарения влаги. 3. Функции и роль подстилки в экосистеме 3.1 Экологическая роль лесных подстилок в миграции техногенных загрязнителей Исследования миграции радиоактивных веществ показали, что после их поступления на поверхность лесной подстилки ...
0 комментариев