ОПИОИДНЫЕ ПЕПТИДЫ И ФИЗИОЛОГО-БИОХИМИЧЕСКИЕ АСПЕКТЫ ИХ ДЕЙСТВИЯ
ОБМЕН РЕГУЛЯТОРНЫХ ПЕПТИДОВ
Механизмы регуляции активности ферментов обмена нейропептидов
ОПИОИДНЫЕ ПЕПТИДЫ ПРИ ВОЗДЕЙСТВИИ СТРЕССОРНЫХ ФАКТОРОВ
ФЕРМЕНТЫ ОБМЕНА НЕЙРОПЕПТИДОВ ПРИ СТРЕССЕ
МАТЕРИАЛЫ ИССЛЕДОВАНИЯ
Метод определения содержания белка
ИССЛЕДОВАНИЕ ВЛИЯНИЯ ОСТРОГО ЭМОЦИОНАЛЬНО-БОЛЕВОГО СТРЕССА НА АКТИВНОСТЬ КПН, ФМСФ-ИНГИБИРУЕМОЙ КП И АПФ
Активность ФМСФ-ингибируемой КП в головном мозге, надпочечниках и семенниках крыс при воздействии острого эмоционально-болевого стресса
Активность КПН в головном мозге, надпочечниках и семенниках крыс при введении лей-энкефалин-арг
Активность АПФ в головном мозге, надпочечниках и семенниках крыс при введении лей-энкефалин-арг
Активность КПН при введении лей-энкефалин-арг на фоне острого ЭБС
Активность АПФ при введении лей-энкефалин-арг у крыс на фоне острого ЭБС
Навигация
Механизмы регуляции активности ферментов обмена нейропептидов
Влияние предшественника лей-энкефалина на активность ферментов обмена регуляторных пептидов головного мозга и периферических органов крыс в норме и при эмоционально-болевом стрессе
222465
знаков
7
таблиц
0
изображений
1.2.2. Механизмы регуляции активности ферментов обмена нейропептидов
Данные о вовлечении ряда ферментов в процессинг нейропептидов, а также о возможности контроля активности ферментов физиологически активными пептидами являются причиной повышенного интереса к исследованию малоизученных механизмов регуляции их активности. Изучение механизмов, по которым осуществляется регуляция активности ферментов, имеет большое практическое значение, так как предполагает управление течением процессов образования или деградации биологически активных пептидов.
Известно, что регуляция активности ферментов процессинга может осуществляться in vivo на разных уровнях [38, 39, 48, 246] :
n на уровне экспрессии гена;
n на уровне процессинга предшественников ферментов;
n на уровне зрелой, активной формы ферментов;
В настоящее время не вызывает сомнений, что активность КПН in vivo может регулироваься на уровне экспрессии гена [38, 202]. Поскольку ген КПН состоит из большого количества регуляторных участков, структура которых различна в разных типах клеток, следовательно, и влияние агентов на экспрессию гена КПН будет различным и избирательным. Так, стимуляция надпочечников инсулином ведет за собой значительное повышение уровня мРНК КПН, изменения в уровне мРНК фермента при этом менее значительны, чем для нейропептида [213, 246]. Тот факт, что влияние различных факторов не всегда согласовано и одинаково для экспрессии генов КПН и экспрессии нейропептидов, позволяет предположить, что регуляция на уровне экспрессии генов не единственный механизм, по которому осуществляется регуляция активности КПН.
Известно, что КПН синтезируется в виде препрофермента, который не обладает ферментативной активностью [38, 39]. Активация неактивной формы фермента происходит в результате отщепления N-концевой последовательности предшественника в секреторных везикулах по мере их продвижения от тел нервных клеток по аксонам, т.е. по мере их созревания [38]. Протеолитическое удаление С-концевой последователь-ности проформы КПН происходит в секреторных везикулах при участии фермента, активируемого ионами Са2+, что приводит к превращению мембраносвязанной формы в растворимую [38, 186, 264]. Теоретически, можно предположить, что регуляция активности КПН на этом этапе возможна за счет изменения концентрации ионов Са2+. Однако тот факт, что в распределении активности между растворимой и мембраносвязанной формами в клетках с различным уровнем экспрессии и активности КПН нет различий [264], говорит в пользу того, что регуляция активности КПН на уровне превращения мембраносвязанной формы в растворимую невозможно [38].Таким образом, доказательств регуляции процессинга КПН в процессе созревания секреторных везикул нет. Что же касается предположения о регуляции активности КПН на уровне превращения неактивного предшественника фермента в активную форму, то оно остается в силе, хотя, на данном этапе еще нет достоверных доказательств этого.
Особый интерес у исследователей вызывает регуляция активности ферментов обмена регуляторных пептидов на уровне зрелой формы. Известно, что изменения активности ферментов достигаются при воздействии ионов металлов, продуктов ферментативных реакций, их субстратов, эндогенных активаторов и ингибиторов, а также изменением рН [38, 39, 48].
Возможность регуляции активности ферментов обмена регуляторных пептидов эндогенными активаторами и ингибиторами пептидной природы описана для вневезикулярных ферментов, в частности для ангиотензинпревращающего фермента [73]. Для внутривезикулярных ферментов (например, КПН) такие данные полностью отсутствуют, поскольку механизмов проникновения в секреторные везикулы веществ пептидной природы не обнаружено.
Обнаружено, что активность внутривезикулярных и вневезикулярных ферментов in vitro ингибируется биологически активными пептидами и их предшественниками [188, 206, 215, 233]. Так показано, что активность КПН in vitro ингибируется ее субстратами - мет5-энкефалин-арг6, лей5-энкефалин-арг6, мет5-энкефалин-лиз6, лей5-энкефалин-лиз6 [119, 193, 194, 205], а также продуктами каталитических реакций фермента – мет- и лей-энкефалином, арг-вазопрессином, веществом Р, тиротропин-рилизинг фактором [119, 193]. Регуляция активности внутривезикулярных ферментов таким способом возможна только в том случае, если при нанесении веществ пептидной природы в секреторных везикулах изменяется суммарная концентрация и нейропептида и его предшественника, что предполагает проникновение субстратов и продуктов каталитических реакций в сформировавшиеся везикулы. Однако установлено, что концентрация активного пептида и пропептида в секреторных везикулах, где осуществляется их процессинг, остается постоянной, что свидетельствует о невозможности регуляции активности ферментов внутривезикулярной локализации таким способом.
В случае вневезикулярных ферментов (например, АПФ) регуляция активности биологически активными пептидами и их предшественниками может иметь место, поскольку фермент является более “доступным” для них. Показано, что активность АПФ in vitro ингибируется энкефалинами, b-эндорфином, веществом Р, АКТГ, брадикинином, b-липотропином [160].
Обнаружено, что in vivo активность КПН и ФМСФ-ингибируемой КП головного мозга, надпочечников и семенников активируется лей- и мет-энкефа-линами, введенными методом инстилляции на конъюнктиву глаза, причем влияние лей-энкефалина более выражено [119]. Механизмы влияния исследо-ванных пептидов на активность ферментов не изучены. Предполагается, что экзогенные лей- и мет-энкефалины влияют на уровень экспресси мРНК КПН и ФМСФ-ингибируемой КП, что приводит к изменению активности фермента.
Сходная тенденция к повышению активности при введении лей- и мет-энкефалинов обнаружена для АПФ [119]. Вероятно, что данные пептиды могут влиять на уровень и активность эндогенных активаторов, способствующих увеличению активность данного фермента.
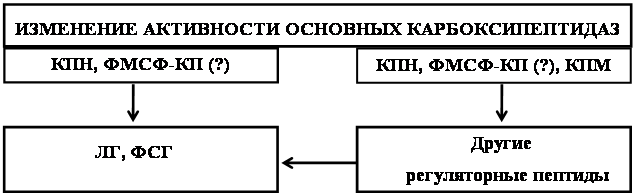
Обнаруженное под воздействием субстратов и продуктов каталитических реакций, изменение активности ферментов обмена регуляторных пептидов, вероятно имеет важный биологический смысл для регуляции уровня нейропептидов при различных патологических состояниях организма, таких как алкоголизм, стресс-реакции и т.д.
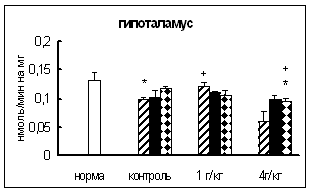
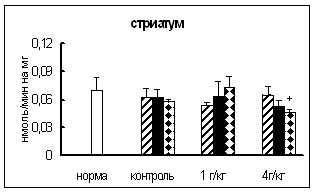
В целом ряде работ имеются данные о повышении активности некоторых ферментов обмена нейропептидов (КПН и АПФ) при введении этанола [19, 41], глюкокортикоидов [47], резерпина, каптоприла [58]. Поскольку in vitro подобных эффектов не обнаружено, то предполагается, что показанное воздействие опосредовано механизмами регуляции активности данных ферментов и является одним из звеньев неспецифической реакции организма на действие экстремальных факторов.
Таким образом, сведения, касающиеся механизмов регуляции активности ферментов обмена регуляторных пептидов активными формами нейропептидов, а также их предшественниками носят фрагментарный характер. Вместе с тем изучение этого вопроса позволит понять принципы функциональной организации ферментов, с тем, чтобы использовать эти сведения на практике при разного рода патологических состояниях организма, таких как алкоголизм, стресс-реакции и т.д.
Похожие работы

... нервной и эндокринной регуляции в организме // Усп. совр. биол. – 1981. – 19, № 3. – С. 380-382. 23. Вернигора А.Н., Бардинова Ж.С., Сметанин В.А., Генгин М.Т. Активность основных карбоксипептидаз в тканях самок крыс на разных стадиях эстрального цикла // Укр. биохим. журн. – 2003. – 75, № 5. – С. 99-102 24. Вернигора А.Н. Карбоксипептидаза Н мозга животных в норме и при действии стресс- ...
0 комментариев