Традиционные биохимические процессы, протекающие с применением дрожжей
Дрожжи в современной биотехнологии
Систематика дрожжей
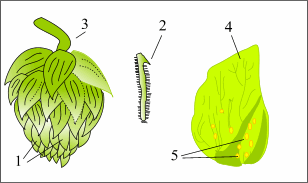
Общая характеристика хмеля
Выделение чистой культуры дрожжевых грибов
Глава Собственные исследования
Выделение дрожжей
Определение видовой принадлежности
Вспомогательные материалы
Навигация
Систематика дрожжей
Выделение чистых культур дрожжевых грибов из шишек хмеля
174948
знаков
8
таблиц
3
изображения
1.4 Систематика дрожжей
Одноклеточная организация дрожжей накладывает столь существенный отпечаток на их внешний облик и на методы работы с ними, что систематика дрожжей долгое время развивалась вполне независимо от систематики мицелиальных грибов. Одно из важных отличий - широкое использование для классификации и идентификации дрожжей физиологических и биохимических признаков. До середины XX в. все одноклеточные грибы рассматривались в качестве достаточно обособленной таксономической группы аскомицетов. Последней точки зрения придерживался, например, В.И.Кудрявцев, автор отечественного определителя дрожжей, который предлагал объединять все дрожжи в самостоятельный порядок Unicellomycetales. В середине XX в. произошло принципиальное событие в систематике дрожжей, когда японскому микологу Исао Банно удалось индуцировать половой цикл размножения у гетероталличных красных дрожжей Rhodotorula glutinis. Полученные им характеристики жизненного цикла однозначно свидетельствовали о принадлежности этих дрожжей к базидиомицетам. Стало очевидным, что среди дрожжей имеются представители совершенно различных таксономических групп грибов, как аскомицетовых, так и базидиомицетовых. После этого большое внимание в систематике дрожжей стало уделяться поиску признаков, позволяющих разделить аскомицетовые и базидиомицетовые виды, даже без наблюдения полного жизненного цикла (так называемых признаков аффинитета). В систематике дрожжей стали активно использоваться биохимические и цитологические признаки.( Хиггинс И., Бест Д., Джонс Д.М.).
Современный период изучения биологического разнообразия характеризуется интенсивным развитием филогенетической систематики, которая направлена на реконструкцию конкретных путей исторического развития организмов. В микробиологии филогенетическая систематика получила мощный импульс развития лишь в самом конце XX в. в связи со сравнительным изучением консервативных нуклеотидных последовательностей в рРНК. К настоящему времени расшифрованы и помещены в компьютерные банки данных, доступные по сети Интернет, нуклеотидные последовательности рРНК у представителей всех известных видов дрожжей. Это позволяет строить филогенетические деревья, отражающие эволюцию их рибосомальных генов.
Оказалось, что группирование дрожжей на основе сходства нуклеотидных последовательностей рРНК во многих случаях не совпадает с группированием по фенотипическим признакам. Многие традиционные признаки, используемые в классификации дрожжей, такие как характеристики вегетативного размножения, форма аскоспор, способность к сбраживанию и ассимиляции сахаров, стали считаться ненадежными, не пригодными для определения филогенетического родства. Секвенирование рРНК (рДНК) сейчас считается необходимым при описании новых видов дрожжей.( Главачек Ф.).
Один из главных вопросов, который активно дискутировался до последнего времени, положение дрожжей в общей системе грибов. Являются ли они предковыми примитивными формами аско- и базидиомицетов, давшими начало более продвинутым и сложно организованным мицелиальным грибам, или вторично упростившимися, возникшими независимо в разных филогенетических линиях грибов в результате конвергенции? Окончательный ответ на этот вопрос был получен лишь недавно в результате развития молекулярно-филогенетической систематики. Сейчас считается окончательно доказанным полифилетическое происхождение дрожжей, их независимое возникновение среди аскомицетовых и базидиомицетовых грибов. После обнаружения базидиомицетовых дрожжей в зимологии возникло представление о дрожжах, как о чисто морфологической, или экоморфологической группе грибов (жизненной форме), лишенной таксономического содержания. В то же время, дрожжи встречаются лишь в некоторых филогенетических линиях грибов, в которых имеются также близкородственные виды, существующие в основном в мицелиальной форме. Примерами могут служить аскомицеты Endomyces, Blastobotrys, базидиомицеты Tilletiopsis, Trichosporonoides, не образующие почкующихся одиночных клеток. Несмотря на отсутствие одноклеточных ассимилятивных стадий такие виды также включают в определители дрожжей, так как филогенетически они очень близки к «настоящим» одноклеточным дрожжам. Поэтому с точки зрения филогенетической систематики, целиком сводить понятие «дрожжи» только к одноклеточной жизненной форме грибов не представляется целесообразным.
Систематика дрожжей, поиск об их места в общей системе грибов продолжают активно развиваться, и в этой области еще не выработано устоявшихся, стабильных представлений. Тем не менее, со времени первых описаний сахаромицетов зимологами пройден очень большой путь в исследовании разнообразия дрожжевых грибов. Основные этапы этого пути отражены в серии определителей голландской зимологической школы, которые выходили с интервалом в 10-20 лет.( Коновалов С.А.).
Понятие вида - одно из сложнейших в биологии. Разработка концепции вида - особенно трудная задача в применении к микроорганизмам, в частности к грибам, по причине их плеоморфизма, сложности жизненных огрпмпциклов, наличия стабильных несовершенных форм, неопределенности границ индивидуума и популяции.
В биологии термин «вид» используется в разных значениях. С одной стороны, о виде говорят как о наименьшей единице классификации, минимальном элементе разнообразия, который мы в состоянии распознать с помощью определенного набора таксономических признаков. Стандартный набор таких признаков, используемых в таксономии дрожжей, постоянно увеличивается, что позволяет выделять все более «мелкие» виды. В результате число известных видов дрожжей за последние десятилетия существенно возросло не только за счет обнаружения новых форм в природе, но и дробления уже известных «крупных», «гетерогенных» видов, характеризующихся значительной изменчивостью. Границы между видами при этом проводятся либо исходя из опыта и интуиции систематика (типологическая концепция вида), либо на основании явного соглашения между систематиками об уровне отличий, достаточном для выделения самостоятельного вида (номиналистическая концепция). В последнем случае вид - это не более чем абстракция, и границы вида устанавливаются исходя из чисто утилитарных представлений об определенном уровне общего сходства между штаммами дрожжей.( Коновалов С.А.).
Наряду с этим в биологии существует и принципиально иное представление о виде, как о целостной, устойчивой и достаточно гетерогенной системе. При этом основное внимание уделяется не столько уровню различий между особями, сколько степени дискретности, отсутствию переходных форм между видами. Такая дискретность наиболее выражена у животных и растений, то есть организмов, размножающихся преимущественно половым путем. Обмен генами при половом размножении как раз и представляет собой механизм, поддерживающий целостность вида, а прекращение такого обмена приводит к возникновению дискретности разнообразия. Поэтому для животных и растений применима биологическая концепция вида: вид - это система популяций, генетически изолированных от популяций других видов и характеризующихся интерфертильностью, то есть способностью особей скрещиваться с образованием плодовитого потомства.
Организмы разных видов не могут скрещиваться, или скрещиваются, но не дают плодовитого потомства. Такая концепция вида применима и ко многим грибам, у которых имеется половой процесс. Изучение интерфертильности играет существенную роль в определении границ видов и у совершенных дрожжей с хорошо выраженной половой стадией в жизненном цикле.
Интерфертильность легко проверяется, если имеются штаммы, представляющие различные типы спаривания гетероталличных дрожжей. Однако гомоталлизм, вегетативное размножение в диплоидной стадии, конъюгация аскоспор в аске - затрудняют применение интерфертильности для разграничения видов дрожжей.( Аркадьева З.А., Безбородов А.М., Блохина И.Н.).
Применение биологической концепции вида к дрожжевым грибам осложняется и тем, что половое размножение у них не является обязательным и не играет такой же существенной роли в формировании разнообразия, как у высших животных и растений. Очевидно, биологическая концепция неприменима к несовершенным дрожжам, размножающимся только бесполым путем, и, возможно, полностью утерявшим половую стадию в ходе эволюции. У таких дрожжей выделение видов неизбежно носит номиналистический характер.
В связи с развитием молекулярно-филогенетической систематики в последнее время изменилось и представление о природе вида у дрожжей. Бурное развитие геносистематики привело к созданию и широкому внедрению новых методов, включая расшифровку и сравнение нуклеотидных последовательностей рДНК, ПЦР-анализ амплифицированной ДНК и др. Это породило надежды на создание абсолютно объективных технологий видовой идентификации. На первый план вышла концепция «элементарных» монофилетических видов, то есть таких групп штаммов, внутри которых полностью отсутствует филогенетическая структура, выявляемая на основании анализа нуклеотидных последовательностей в наиболее консервативных генах. Самостоятельность многих видов дрожжей стала активно пересматриваться с молекулярно-генетических позиций. Это вызвало определенное «высокомерие» по отношению к классическим представлениям о смысле выделения видов и видовой формы организации вообще. В связи со сложностью и дороговизной молекулярно-генетических методов возникла проблема практической недостижимости «правильной» видовой идентификации при проведении исследований, требующих массового определения большого числа штаммов, например, экологических, инвентаризационных, биоресурсных. Вместе с тем, на фоне сохраняющегося пиетета перед молекулярно-генетическими методами в последнее время появляется все больше свидетельств их недостатков, спорности некоторых положений, противоречий с эволюционно-филогенетическими концепциями. Иными словами, возникла проблема соотношения новейших молекулярных подходов к выделению видов с традиционными, основанными на морфологических и физиологических признаках. Решение этой проблемы должно определить развитие систематики дрожжей в ближайшем будущем.( Коновалов С.А.).
Набор признаков, на основании которых выделяются виды у дрожжей, постоянно меняется в связи с принятием той или иной концепции вида, в результате пересмотра таксономической значимости признаков, с появлением новых методов и технологий. Систематика грибов до последнего времени основывалась главным образом на морфологических признаках, таких как пигментация, строение конидиеносцев, форма спор и т.п. Одноклеточные грибы не обладают такой сложной разнообразной морфологией, как мицелиальные. Поэтому уже с первых работ Э.Хансена и А. Клюйвера в 20-х гг. XX в. разделение видов дрожжей основывалось не только на морфологических, но и на физиологических признаках, таких как способность к сбраживанию и аэробной ассимиляции различных углеводов. Количество используемых для описания вида источников углерода и азота было значительно расширено в 50-е гг. Викерхэмом, который также разработал стандартные среды для постановки физиологических тестов. Эти критерии прочно вошли в практику классификации и идентификации дрожжей и используются до настоящего времени.
По своей природе признаки, используемые в систематике дрожжей, можно разбить на следующие группы.
Макроморфологические (культуральные) признаки, которые характеризуют рост штамма на различных средах. К ним относятся особенности роста в жидких средах (образование пленки, мути, осадка), формирование гигантской колонии и ее характеристики, образование плодовых тел.( Работнова И. Л).
Микроморфологические признаки - особенности клеточной морфологии: размеры и форма клетки, тип вегетативного размножения, строение мицелия и псевдомицелия, формирование бесполых спор (баллистоспор, хламидоспор), характеристики полового размножения (гологамия или педогамия, форма аскоспор и др.
Цитологические признаки - особенности строения клеток и клеточных структур, например, строение клеточной стенки, структура септ мицелия.
Физиологические признаки, определяющие тип питания и способность к росту в различных условиях. К ним относится определение способности к анаэробному сбраживанию или аэробной ассимиляции различных источников углерода, способность к росту при различных значениях pH, осмотического давления среды, устойчивость к различным ингибиторам роста.( Шлегель Г).
Биохимические признаки, характеризующие химический состав клетки и отдельных ее компонентов: образование ферментов, специфических метаболитов, внеклеточных продуктов, например, моносахаридный состав внеклеточных полисахаридов.
Генетические признаки, включающие характеристики генома.
Арсенал генетических признаков и методов их определения особенно расширился в последнее время в связи с интенсивным развитием молекулярной биологии и стал играть ключевую роль в систематике дрожжей. К ним относятся нуклеотидный состав ДНК, степень гомологии ДНК у разных видов, наличие уникальных олигонуклеотидных последовательностей в геноме, последовательность нуклеотидов в определенных генах.
Экологические признаки, определяющие характер распространения вида в природных местообитаниях, а также чувствительность или резистентность к различным экологическим факторам, патогенные свойства.
Для выявления всех этих признаков разработаны стандартные методы, описываемые в определителях или специальных руководствах.
Морфологические характеристики, имеющие наибольшее значение для дифференциации родов дрожжей, были подробно рассмотрены выше. Для разделения видов у дрожжей традиционно использовались в основном физиологические характеристики - способность к росту на различных источниках углерода и азота.
Некоторые цитологические и биохимические критерии, разработанные в последние десятилетия, сыграли особенно большую роль в классификации дрожжей на родовом и надродовом уровне, значительно изменив представления об их группировании и филогении. Эти признаки имеет смысл рассмотреть более подробно.
Изучение полисахаридов, которые составляют клеточную стенку и капсулу, оказало существенное воздействие на систематику и преставления о филогении дрожжей. Характеристика моносахаридного состава полисахаридов клеточных стенок используется главным образом для дифференциации дрожжей на родовом и надродовом уровне. В то же время, детальные исследования химического состава клеточной стенки были проведены лишь у небольшого числа видов дрожжей. Наиболее подробно исследовано строение клеточной стенки Saccharomyces cerevisiae и нескольких близких аскомицетовых видов.( Гриневич. А.Г., Босенко. А.М).
Как уже отмечалось, аскомицетовые почкующиеся дрожжи содержат в качестве главного структурного компонента клеточной стенки β (1>3)-глюкан. Другой существенный компонент - комплекс белка и глюкоманнана. Оказалось, что боковые цепи этих полисахаридов, особенно маннана, существенно варьируют по моносахаридному составу у разных видов дрожжей. Моно- и олигосахариды, образовавшиеся при кислотном гидролизе углеводов клеточной стенки, можно разделить с помощью гельфильтрации.
Другой способ, применявшийся для характеристики полисахаридов - сравнение спектров протонного магнитного резонанса, который позволяет определять пропорции и размер боковых цепей в различных маннанах. Считается, что сходные спектры протонного магнитного резонанса клеточных маннанов указывают на близкое родство видов. Эти характеристики оказались очень полезными для классификации дрожжей, например, для группирования родственных видов в многовидовых сборных анаморфных родах типа Candida.
Наряду с глюканом и маннаном почкующиеся аскомицетовые дрожжи содержат около 1-2 % хитина, который почти полностью локализован в областях шрамов почкования. Однако небольшое количество хитина (около 0.1 %) рассеяно по всей клеточной стенке.( Грачёва. И.М.).
Напротив, дрожжи базидиомицетового аффинитета характеризуются намного более высоким содержанием хитина (до 10 %). Таким образом, это различие можно использовать в качестве таксономического признака для разделения аскомицетовых и базидиомицетовых анаморф. Кроме того, оказалось, что содержание хитина в стенках некоторых мицелиальных аскомицетовых дрожжей, например, у видов Saccharomycopsis, значительно выше, чем у истинных одноклеточных дрожжей, таких как Saccharomyces cerevisiae.( http://soil.msu.ru/soilyeasts/pics/AscosporeForms.htm).
Наиболее отличным по моносахаридному составу оказался состав клеточной стенки делящихся дрожжей Schizosaccharomyces. Виды этого рода содержат в дополнение к β-глюканам другой главный структурный полисахарид, а именно α(1>3)-глюкан. Маннан Schizosaccharomyces также отличается по строению от маннана почкующихся дрожжей, главным образом присутствием галактозных остатков. Это позволило предположить, что делящиеся дрожжи имеют уникальный состав клеточной стенки и не родственны другим группам аскомицетовых дрожжей. Позднее это предположение было подтверждено данными по сравнению нуклеотидных последовательностей рРНК.
Еще большее значение играют характеристики моносахаридного состава клеточных стенок в систематике базидиомицетовых дрожжей. Особенно ценным считается такой признак, как присутствие или отсутствие D-ксилозы в клеточной стенке или в экстрактах целых клеток. По этому признаку все гетеробазидиомицетовые дрожжи были разбиты на две группы: спороболомицетовые (отсутствует ксилоза и часто содержится фукоза), которые включают виды родов Rhodotorula, Rhodosporidium, Sporobolomyces, и филобазидиевые (содержат ксилозу и обычно глюкуроновую кислоту), включающие роды Bullera, Cryptococcus, Filobasidium, Phaffia и некоторые виды Trichosporon. Такое разделение также подтверждается другими молекулярно-биологическими критериями.
Несмотря на несомненную важность такого признака, как состав углеводов клеточной стенки, следует подчеркнуть, что он не является решающим критерием в систематике дрожжей, а ценен только в комбинации с другими хемотаксономическими и молекулярными признаками, включая анализ последовательности рРНК. Такой полифазный подход для создания надежной таксономической системы поддерживается большинством систематиков.( Бабицкая В.Г., Стахеев И.В.).
Дрожжи и дрожжеподобные грибы содержат в качестве одного из компонентов цепи переноса электронов кофермент Q, или убихинон.
Убихиноны - группа соединений, в которых к 2,3-диметоксил-5-метилбензоксихинону присоединена в 6 положении боковая цепь из нескольких изопреноидных остатков. У известных гомологов кофермента Q количество изопреноидных остатков варьирует от 5 до 10. По количеству изопреноидных остатков различают до шести типов кофермента Q от Q-5 до Q-10. У базидиомицетовых дрожжей обнаружен также дигидрогенированный гомолог кофермента Q с насыщенной двойной связью в изопреноидных остатках, обозначаемый как Q-10 (H2). Кофермент Q выделяют с помощью тонкослойной хроматографии из гексанового эктракта гидролизованных клеток дрожжей. Для определения типа кофермента Q используют различные хроматографические методы, включая жидкостную, тонкослойную и бумажную хроматографию, а также масспектрометрию.
Тип кофермента Q оказался очень полезным признаком для классификации дрожжей и дрожжеподобных грибов. Прежде всего, он различен у аскомицетовых и базидиомицетовых дрожжей. У первых преобладают убихиноны с 5-7 изопреноидными остатками (Q-5 - Q-7), у вторых - с 8-10 (Q-8 - Q-10). Однако, имеются и исключения, например, убихинон Q-10 обнаружен у представителей родов Lipomyces и Schizosaccharomyces. Особенно важную роль сыграл тип кофермента Q в классификации анаморфных дрожжей на родовом уровне. Считается, что этот признак не должен существенно варьировать внутри рода, и отличия по типу кофермента Q достаточно для отнесения видов к разным родам в том случае, когда это сопровождается и другими существенными различиями, например, по морфологическим признакам.
Определение типа кофермента Q, вместе с изучением моносахаридного состава клеточных стенок сыграло решающую роль в переклассификации родов анаморфных баллистоспоровых дрожжей (Bensingtonia, Bullera, Sporobolomyces, Udeniomyces), дрожжей, характеризующихся образованием почек на стеригмах (Sterigmatomyces, Fellomyces, Kurtzmanomyces). Все эти роды характеризуются одним типом кофермента Q. В то же время, в таких родах, как Cryptococcus, Candida, тип кофермента Q варьирует по видам, что подтверждает их условность и филогенетическую гетерогенность.
Развитие методов электронной микроскопии позволило использовать для классификации дрожжей ряд цитологических признаков. Выше уже были рассмотрены такие характеристики, как ультраструктура клеточной стенки, цитологические особенности образования почки (голобластическое и энтеробластическое почкование), которые оказались различными у аскомицетовых и базидиомицетовых дрожжей. Еще одним важным цитологическим признаком является ультраструктура септ мицелия у диморфных дрожжеподобных грибов. Деление клеток мицелия начинается с образования тонкого кольца на клеточной мембране. Кольцо начинает центростремительно расти и разделяет клетку. Затем на внешней поверхности мембраны откладывается вновь синтезируемый материал клеточной стенки, формируя септу.
Детальные исследования, проведенные с помощью электронной микроскопии, показали, что ультраструктура этих септ существенно различается у разных групп грибов и может служить хорошим критерием для их филогенетической классификации.
В гифах аскомицетовых дрожжеподобных грибов септы в основном гомогенные и электроннопрозрачные. В септах имеются поры, которые достаточно велики для прохода ядер. С обеих сторон поры часто располагаются мелкие мембранные пузырьки, так называемые тельца Воронина. Гифы дрожжеподобных грибов рода Ambrosiozyma имеют септы с сильно утолщенным центральным участком. У некоторых дрожжеподобных грибов в центре сформированной септы имеется лишь очень узкий мембранный канал, так называемая микропора. У других видов в септе могут формироваться множественные каналы - плазмодесмы. Наличие таких плазмодесм явилось важным свидетельством о наличии филогенетической связи анаморфного рода Zygozyma с семействами Lipomycetaceae и Dipodascaceae.( Гречушкина Н.).
У большинства базидиомицетовых дрожжей в центре септы формируется пора, имеющая сложное строение: края септы раздуты в виде тора, а с двух сторон поры имеются характерные мембранные образования - парентосомы. В некоторых исследованиях было показано, что тороидально раздувшиеся септы (долипоры) на самом деле представляют собой артефакт химической фиксации, используемой при подготовке образцов к электронной микроскопии. Такие бочкообразные вздутия отсутствовали в образцах, подготовленных с помощью быстрого замораживания. Тем не менее, эти артефакты четко воспроизводятся у одних и тех же видов, коррелируют с другими таксономическими признаками и поэтому могут использоваться в систематике. Тонкое строение комплекса долипор и парентосом служит важным диагностическим признаком для классификации базидиомицетовых дрожжеподобных грибов.
Аскомицетовые и базидиомицетовые грибы легко отличить по характеру полового размножения: формированию эндогенных спор в асках у аскомицетов и экзогенных спор на базидиях у базидиомицетов. Аналогичные структуры формируют при половом размножении и дрожжевые грибы. Поэтому особую сложность для классификации представляют анаморфные дрожжи, у которых отсутствует половая стадия в жизненном цикле. Формально такие грибы относят к особому классу Deuteromycetes, однако по сути они представляют собой лишь стадии в полном жизненном цикле аскомицетов или базидиомицетов. Половое размножение у таких дрожжей может отсутствовать по разным причинам. Во-первых, многие дрожжи гетероталличны, и для осуществления полового процесса необходимы штаммы разных типов спаривания. В чистой культуре таких дрожжей, представленной только одним типом спаривания, половое размножение невозможно. Во-вторых, половой процесс может запускаться лишь в определенных условиях, например, при наличии определенных химических факторов, которые могут отсутствовать в лабораторной среде. Наконец, у многих видов способность к половому размножению, по-видимому, вообще утеряна в ходе эволюции.( Грачёва. И.М.).
По совокупности морфологических и физиологических признаков, которые использовались на первых этапах развития систематики дрожжей, отличить аскомицетовые и базидиомицетовые дрожжи в анаморфном состоянии было практически невозможно. Это привело к тому, что некоторые крупные роды несовершенных дрожжей включали анаморфы как аскомицетов, так и базидиомицетов. В первую очередь это относится к роду Candida, описанному в 1923 г. Диагноз рода был очень расплывчатым: «Немногочисленные гифы, стелющиеся, распадающиеся на короткие и длинные фрагменты. Конидии, возникающие путем почкования из гиф или на вершине одна другой, мелкие и бесцветные». Под такое описание подходили самые различные дрожжеподобные грибы, и поэтому в дальнейшем оказалось возможным включение в этот род многих видов, явно неродственных друг другу. К 1970 г., в котором был выпущен полный определитель дрожжей дельфтской школы, количество видов, включенных в род Candida, возросло до 81, и он стал самым многовидовым родом среди дрожжей.( Азов а Л. Г.).
Второй по числу видов дрожжевой род Torulopsis существовал с 1895 г. и объединял аспорогенные дрожжи, не соответствующие описанию рода Candida только по одному признаку - отсутствию способности к образованию субстратного септированного или псевдомицелия. При росте на плотных средах мицелиальность настолько меняет облик колонии дрожжеподобных грибов, что на начальном этапе систематики дрожжей, когда основной упор делался именно на морфологические характеристики, этому признаку безусловно был придан статус родового. Однако с увеличением разнообразия описанных видов стало ясно, что способность к образованию псевдомицелия крайне ненадежный, сильно варьирующий в зависимости от штамма и от условий культивирования признак, имеющий низкую таксономическую ценность. В связи с этим в 1978 г. было предложено объединить роды Candida и Torulopsis в один род. Это было осуществлено в следующем издании определителя дрожжей, где был представлен единый род Candida, включающий 196 видов и заведомо полифилетический. Наиболее вескими аргументами полифилетичности рода были обнаружения совершенных стадий для некоторых видов Candida, которые, как оказалось, соответствовали различным родам известных аскоспоровых дрожжей. Такие пары анаморфа - телеоморфа включали, например Candida famata - Debaryomyces hansenii, Candida pulcherrima - Metschnikowia pulcherrima, Candida robusta - Saccharomyces cerevisiae и др. В 1966 г. новозеландская исследовательница Ди Менна описала три новых вида Candida gelida, Candida nivalis, Candida frigida, выделенные ею из антарктических почв. Всего несколько лет спустя у этих дрожжей также были обнаружены совершенные стадии, которые свидетельствовали об их принадлежности к базидиальным грибам. Таким образом, род Candida оказался группой несовершенных дрожжей, объединяющей анаморфы как аскомицетов, так и базидиомицетов.( И.М. Грачёва, Л.А. Иванова. В.М. Кантере.).
После доказательства полифилетической природы таких крупных дрожжевых родов, как Candida, в зимологии начался активный поиск признаков, которые могли бы дифференцировать анаморфы аско- и базидиомицетов. К настоящему времени в систематике дрожжей используется целый набор таких признаков аффинитета, благодаря которым все дрожжи удается четко разбить на две группы - аскомицетовые и базидиомицетовые, независимо от телеоморфного или анаморфного состояния культуры.
Развитие методов секвенирования рРНК окончательно решило проблему определения аффинитета несовершенных дрожжей. Одно из главных преимуществ систематики, основанной на сравнении нуклеотидных последовательностей консервативных генов - возможность классификации на одной и той же основе как совершенных видов дрожжей, обладающих полным жизненным циклом, так и их анаморф.( Розманова Н.В., Бочарова Н.Н.).
Вновь описываемые виды должны типифицироваться штаммами, которые помещают в крупные коллекции, где они поддерживаются в живом состоянии. Это делает их доступными для научной общественности. Кроме хранилищ таких штаммов коллекции выполняют различные функции: научные, учебные, производственные. Они проводят патентование практически ценных штаммов, осуществляют обмен и выдачу культур для разных целей, проводят таксономические исследования, составляют периодически публикуемые каталоги. Коллекции дрожжей различаются как по своему объему, так и по направленности. Есть очень крупные, хорошо известные в научном мире коллекции, где проводятся широкие исследования по систематике дрожжей, разрабатываются способы наилучшего хранения штаммов. Есть и небольшие, но очень ценные коллекции, где собраны штаммы с конкретной целью, служащие для выполнения специальных исследований.( Жвирблянская А.Ю., Исаева В.С.).
Крупнейшая коллекция дрожжей - CBS (Centraalbureau voor Schimmelcultures). Эта коллекция была создана в 1904 г. по решению 11-го международного Ботанического Конгресса в Вене. Она поддерживается Королевской Нидерландской Академией Искусств и Наук и до 2000 г. располагалась на родине Левенгука в г. Дельфте. В настоящее время коллекция CBS находится в г.Утрехте и является мировым центром по изучению систематики дрожжей. Главным образом на базе этой коллекции была создана серия определителей дрожжей, содержащих описания всех известных видов и диагностические ключи для их идентификации.( Бабьева И.П., Голубев В. И.).
Среди других крупных коллекций дрожжей ВКМ (Всероссийская коллекция микроорганизмов) и ВКПМ (Всероссийская коллекция промышленных микроорганизмов) в России, ATCC (American Type Culture Collection) в США, IFO (Institute of Fermentation in Osaka) в Японии, CCY (Culture Collection of Yeasts) в Словакии, NCYC (National Collection of Yeast Cultures of the United Kingdom) в Англии. Существует также ряд крупных специализированных коллекций дрожжей, например, генетически модифицированных штаммов Saccharomyces cerevisiae, штаммов, используемых в виноделии и других биотехнологических процессах.
Основная проблема, с которой сталкиваются работники микробиологических коллекций - необходимость длительного поддержания чистых культур в жизнеспособном состоянии. Для хранения дрожжевых культур используются разные методы, каждый из которых имеет свои преимущества и недостатки.( Бабицкая В.Г., Стахеев И.В.).
Наиболее доступный и широко применяемый метод хранения - поддержание культур путем их периодических пересевов, обычно в пробирках со скошенным агаром. Сроки пересевов определяются скоростью высыхания среды и зависят от температуры и влажности помещения. Промежутки между пересевами можно увеличить за счет более плотного закупоривания пробирок и снижения температуры хранения. Хранение в холодильной камере при температуре около 5°C дает возможность увеличить сроки пересевов до 2-3 лет. Для предотвращения высыхания культур используют заливку культур минеральным маслом. Этот способ позволяет сохранять культуры большинства видов дрожжей без пересевов в течение 10 и более лет.( Аркадьева З.А.).
Другой способ, используемый для длительного хранения дрожжевых культур - лиофилизация, то есть высушивание под вакуумом из замороженного состояния. Лиофилизированные культуры хранят в запаянных стеклянных ампулах при комнатной температуре или в холодильнике. Такие культуры могут сохраняться в жизнеспособном состоянии в течение нескольких десятилетий.
Недостатком этого способа является невозможность визуального контроля за жизнеспособностью культуры. Кроме того, не все виды дрожжей выдерживают процесс лиофилизации, а при длительном хранении лиофилизированных культур могут происходит существенные изменения в их метаболизме.( Хиггинс И., Бест Д., Джонс Д.М).
В последнее время все шире применяется еще один способ длительного хранения культур микроорганизмов - замораживание в жидком азоте. Замороженные в ампулах культуры хранят в специальных контейнерах-рефрижераторах с жидким азотом при температуре -196°C. Такой способ позволяет сохранять жизнеспособные культуры дрожжей в течение практически неограниченного времени.( Безбородов А.М.).
Определение видовой принадлежности дрожжей - сложная процедура, требующая не только большого опыта, но и, как правило, достаточно длительных лабораторных исследований, связанных с постановкой серии тестов для определения рассмотренных выше морфологических, физиологических и биохимических признаков. Лишь после составления достаточно полного описания штамма возможна его надежная видовая идентификация. В последнее время для идентификации дрожжей по таким описаниям, наряду с традиционными дихотомическими ключами, широко используются компьютерные технологии нумерической идентификации. Такая идентификация может осуществляться на основе любой СУБД (средство управления базами данных), в которую внесены стандартизованные признаки всех видов дрожжей. В процессе идентификации программа поочередно сравнивает описание идентифицируемого штамма с описанием каждого вида и высчитывает уровень сходства (долю совпавших признаков от общего количества использованных признаков). Результирующая информация представляет собой список видов, ранжированный по уровню сходства с идентифицируемым штаммом. Существуют специализированные программы для идентификации дрожжей. (Розманова Н.В., Бочарова Н.Н.).
Некоторые коллекции дрожжей предоставляют возможность поиска наиболее сходных штаммов с использованием Интернета. В последние годы бурно развивается система генобанков, доступных также через Интернет, позволяющих искать наиболее сходные нуклеотидные последовательности. Однако следует помнить, что при идентификации с использованием таких формальных процедур возможны серьезные ошибки, поэтому для надежного определения вида дрожжей необходима консультация со специалистом-систематиком, имеющим большой опыт изучения разнообразия этих организмов. В первую очередь это относится к идентификации штаммов, выделенных из природных местообитаний.
Похожие работы
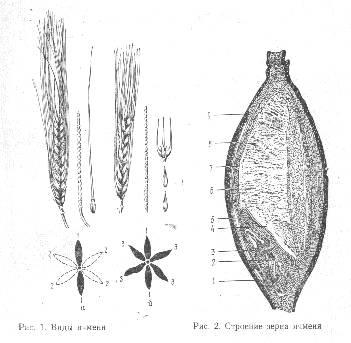
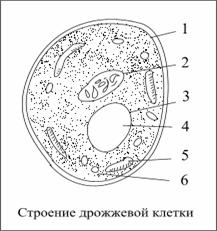
... повышенной температуре, несвоевременное шпунтование танков при дображивании, приготовление заторов из сильно растворенных солодов, отрицательно влияют на пенообразование. Раздел 3.Технологический расчет сырья для производства пива Наименование Условное обозначение Количество на 100 кг зернового сырья Объем сусла Vc 54,51583365 Горячее сусло Vгс 56,696467 Холодное сусло Vхс 53, ...
... этих препаратов превосходит активность ферментов солода по осахаривающей способности в 3-4 раза, по разжижающей - в 8-10 раз, по декстринирующей - в 10-20, по протеолитической - в 15-20 раз. Опыт использования в пивоваренной промышленности МЭК свидетельствует о том, что создание широкого набора эффективных мультиэнзимных композиций способствует успешной переработке повышенных количеств ...
... любого, кто украдет росток, чтобы его возделывали в другой стране.[11] Учитывая все вышесказанное, утверждать с полной уверенностью - так сказать, однозначно, кто именно и когда первым стал использовать в технологии пивоварения хмель, весьма затруднительно. Но это, впрочем, не так уж важно. Главное то, что в результате у пива появился тот "пивной" вкус, который мы хорошо знаем сегодня.[11] Что ...
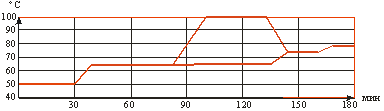
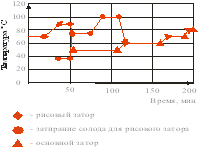
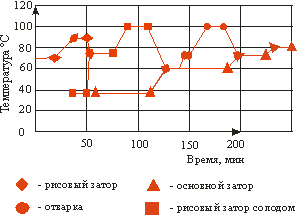
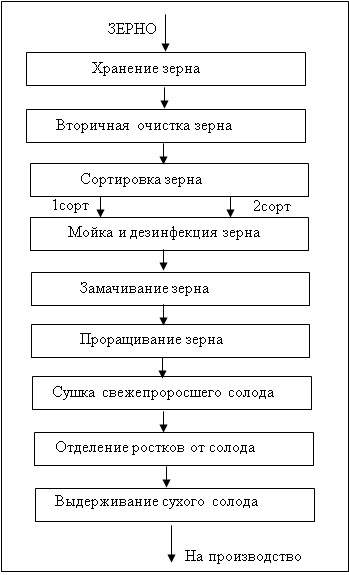
... со 100 до 138°С остается неизменным. При дальнейшем повышении температуры (до 143°С) уровень аминокислот падает, что связано с усилением реакции меланоидинообразования. 2. ТЕХНОЛОГИЧЕСКАЯ ЧАСТЬ 2.1 Описание технологии производства пива «Рецептура №1», «Рецептура №2» и «Рецептура №3» В технологической схеме производства пива можно выделить несколько этапов (приложение 1): подготовка воды ...
0 комментариев