Навигация
Физические вопросы строения и функционирования биологических мембран
33779
знаков
0
таблиц
2
изображения
Физические вопросы строения и функционирования биологических мембран
Содержание
1. Физические вопросы строения и функционирования биологических мембран
1.1 Пассивный и активный транспорт веществ через мембранные структуры
1.2 Транспорт неэлектролитов путем простой и облегченной диффузии
1.3 Явление переноса. Общее уравнение переноса
1.4 Диффузия. Пассивный перенос неэлектолитов через биомембраны, уравнение Рика. Транспорт неэлектролитов через мембраны путем простой и облегченной (в комплексе с переносчиком) диффузии
1.5 Молекулярный механизм активного транспорта ионов
1.6 Проницаемость
Список использованных источников
1. Физические вопросы строения и функционирования биологических мембран
Структурной и функциональной единицей живого организма является клетка. Клетка обменивается с окружающей средой и веществом, и энергией, и информацией. Энергия питательных веществ, поступающих в клетку, расходуется на выполнение ею разнообразных функций.
Все функции клетки тесно связаны с ее структурой.
Напомним в общих чертах, что любая клетка состоит из окруженной плазматической (клеточной) мембраной цитоплазмы, в которой находятся ядро клетки, органеллы и различные включения. К органеллам клетки относятся митохондрии, лизосомы, рибосомы, аппарат Гольджи, эндоплазматический ретикулум. Все они, в том числе и ядро клетки, тоже имеют мембраны, и основная цель данного параграфа состоит в ознакомлении с современными представлениями о структуре и функциях биологических мембран.
Роль мембран в строении и функционировании клетки чрезвычайно велика. Как заметил Бернал Д. “только после образования мембраны вокруг клетки мы действительно имеем то, что с полным правом может быть названо организмом”. Именно через мембраны происходит обмен веществ в организме. Нарушение структуры мембран ведет к нарушению их функции и, следовательно, к нарушению функциональных состояний организма в целом. Нарушение функций биологических мембран является причиной многих заболеваний.
Несмотря на большое разнообразие клеток и организмов в природе, строение и многие функции биологических мембран во многом одинаковы, что и позволяет нам рассматривать их основные свойства безотносительно к виду клеток.
Строение биологических мембран изучается уже более 80 лет. Еще в 1902 году Овертоном была выдвинута первая модель мембраны в виде тонкого слоя липидов. С тех пор представления о строении мембран постоянно усложнялись, совершенствовались, дополнялись и к настоящему времени они существенно отличаются от той первой простейшей модели Овертона. Однако и по современным представлениям основу, матрицу любой биологической мембраны составляет липидный бислой. Каковы же физические свойства липидов и липидного слоя?
Мембранные липиды - это низкомолекулярные вещества, близкие по своим свойствам и жирам. Характерная особенность любой липидной молекулы состоит в том, что она построена из двух физически разных частей: из головки, составляющей примерно четверть длины молекулы и двух длинных неполярных хвостов. Хвосты представляют собой длинные цепи жирных кислот, которые могут быть как насыщенными, так и ненасыщенными. Головки липидов тоже могут иметь разное строение, но для липидов биомембран наиболее характерны производные сахаров и фосфорной кислоты - в соответствии с этим различают глико- и фосфо-липиды.
Головки липидов либо заряжены отрицательно, либо электрически нейтральны, но имеют неравный нулю дипольный момент. Положительно заряженных головок нет и это играет очень важную роль в формировании всего электрического заряда мембраны и в ее функционировании. Поскольку головки липидов полярны, то они хорошо взаимодействуют с полярными растворителями, в частности с водой, поэтому головки называют гидрофильной частью липида. Хвосты, наоборот, не взаимодействуют с водой, - они гидрофобны, но они хорошо взаимодействуют с неполярными веществами и растворителями.
Эти свойства липидов приводят к тому, что они одинаково плохо растворяются и в полярных растворителях (вода) - мешает хвост, и в неполярных (масло) - мешает головка. Если липиды поместить на поверхность воды, то они все станут на “голову” - и вверх хвостами. В масле же картина будет обратной.
Так как вода является основным, универсальным растворителем в биологических системах (цитоплазма, например, на 95% состоит из воды), то нас интересует, как будут вести себя липиды, помещенные “внутрь” воды.
Оказывается, что в этом случае они тоже стремятся быть всегда направленными полярными головками в сторону воды, а неполярными хвостами - внутрь.
При этом могут образовываться различные пространственные структуры липидов. При малых концентрациях липидов сначала образуются мицеллы, а при более высокой концентрации образуются липосомы, или везикулы ”пузырьки”. Наконец, при некоторой концентрации липидов мицеллы слипаются и образуют плоский бимолекулярный слой - липидный бислой.
Толщина такого липидного бислоя не превышает 6¸7 нм. Липидные бислои достаточной протяженности стремятся замкнуться сами на себя, чтобы лучше спрятать гидрофобные участки от воды. Так происходит самопроизвольный процесс сборки мембраны, то есть их “самосборка”.
Впоследствии было показано, что липидный бислой мембран окружен с обеих сторон из фибриллярных (нитевидных) белков, а снаружи их - еще и слоем глобулярных белков. Кроме липидов и белков в составе мембран много холестерина. Общая толщина такого “бутерброда” составляет 8¸9 нм: сверху и снизу два “ломтя” белка, а внутри “масло” - липиды. Внутри мембраны возможны поры, общая площадь которых обычно не превышает 1% от всей площади мембраны.
Представленная модель объясняет многие свойства мембран - их эластичность, избирательную проницаемость (хорошую для неполярных, то есть гидрофобных, соединений и плохую для полярных), в связи с чем она долгое время была принята в качестве унитарной, то есть единой модели всех биологических мембран. Однако последние данные свидетельствуют о том, что на самом деле все обстоит несколько сложнее.
В липидном бислое мембран может быть много белков, встроенных в сам липидный бислой, которые пронизывают липидный бислой насквозь.
По словам одного из авторов этой модели, названной жидкостно-мозаичной, мембрана представляется ему в виде липидного моря, в котором плавают белковые айсберги.
Соотношение между количеством белков и количеством липидов в мембране неодинаково и зависит от функционального назначения клетки.
Так, в мембране эритроцитов 75% площади занимают липиды, а 25% - белки. Вместе с тем весовое соотношение почти обратное (так как белки значительно тяжелее липидов) - в этой же мембране по весу белков в 2,5 раза больше, чем липидов. В мембране некоторых бактерий содержится белков в 5¸6 раз больше, чем липидов, а в миелиновой мембране, наоборот - белка в 2,6 раза меньше, чем липидов.
Белки, входящие в состав мембран, обычно делят по их положению в мембране на периферические и интегральные (проникающие в мембрану), а по их функциональным характеристикам - на структурные и ферментативные (катализирующие).
Роль белков в функционировании мембраны чрезвычайно велика. Структурные белки участвуют в построении мембран вместе с липидами, взаимодействуют стехиометрически с другими белками, участвуют в транспорте веществ и т.п. Не меньшее, если не большее значение имеют и ферментативные белки, главными их которых являются АТФ - азы (аденозинтрифосфатазы). Кроме них в мембранах отдельных клеток могут находиться и другие ферменты: флавины, питохромы, дегидрогеназы и другие, которые принимают активное участие в метаболизме клеток.
В настоящее время большой интерес представляет изучение информации белков (то есть изменение их пространственной формы, структуры), поскольку предполагается, что такие жизненно важные процессы, как окислительное фосфорилирование, активный транспорт веществ, химические реакции при фотосинтезе, проведение нервного импульса и др. тесно связаны с изменением пространственной укладки атомов, то есть с конформацией полипептидной цепи белковых молекул.
Роль липидов в мембранах тоже не сводится лишь к приданию мембранам определенной физической структуры и низкой проницаемости для полярных веществ. Сейчас установлено, что физическое состояние, главным образом вязкость липидного бислоя самым непосредственным образом влияет на каталитическую активность мембранных ферментов, на проницаемость мембран, а значит, на процессы обмена веществ в клетках.
Чем меньше вязкость липидов, тем обычно лучше идут процессы обмена веществ в клетке. Методом ЭПР - спектроскопии было установлено, что микровязкость липидного слоя в мембране эритроцитов, митохондрий, нервных волокон составляет 30¸100 м Па, то есть близка к вязкости подсолнечного масла. Это свидетельствует о том, что липидный слой находится в жидком состоянии.
При многих видах патологий, а также при воздействии биологически активных соединений вязкость мембран изменяется.
Чем выше подвижность хвостов фосфолипидов, тем меньше вязкость мембран, и тем лучше их проницаемость для диффундирующих веществ. Поэтому вязкость бислоя очень важна для всей функциональной деятельности ферментов, находящихся в липидном слое и других.
Вязкость мембраны сильно зависит от агрегатного состояния бислоя (жидкое и твердое), то есть от температуры.
В твердом липидном бислое хвосты фосфолипидов имеют трансконформацию, а в жидком бислое возможны тепловые переходы из транс-конформации в гош-конформацию (транс - гош - переходы).
Рассматривая жидкостно-мозаичную модель мембран, надо всегда иметь в виду, что отдельные молекулы липидного бислоя не “сидят” на одном месте, а непрерывно меняются местами, перемещаясь вдоль слоя, то есть вдоль мембраны с внушительной скоростью в 5 мкм/с. Участвуют в этом движении и белки. Это явление хаотического перемещения молекул липидов и белков вдоль поверхности мембраны называется латеральной (lateral - боковой) диффузией. Скорость латеральной диффузии белков значительно меньше, чем у липидов, кроме того, часть пронизывающих мембрану белков оказывается “заякоренной” на внутриклеточные белки и в латеральной диффузии не участвуют.
Кроме латеральной диффузии, возможен также переход липидной молекулы из одного монослоя в другой - этот процесс называют флип-флопом. Однако вероятность флип-флопа в 1010 раз меньше, чем латеральной диффузии.
Мембранные белки и липиды помимо поступательного движения участвуют и во вращательном движении, или, как говорят, - во вращательной диффузии. При этом угловая скорость вращения белков и липидов весьма велика. Например, при нормальных температурах она составляет:
- для фосфолипидов - 109 рад/с;
- -для родопсина - 106 рад/с;
- для цитохромоксидазы - 104 рад/с.
Ассиметрия мембран проявляется в том, что внутренние и наружные по отношению к клетке стороны любой биологической мембраны всегда имеют разный липидный и белковый состав. Молекулы углеводов располагаются только на внешней стороне мембраны. Ассиметричная ориентация ферментативных и транспортных белков в мембране приводит к наличию преимущественно направления активного транспорта веществ через мембрану, что играет исключительно важную роль для функционирования клетки в целом.
Похожие работы
... материалы хорошо описываются в рамках квантово-механической фононной Модели строения и функционирования клеточных мембран, что позволяет утверждать: “ФОНОН – КВАНТ биологической (клеточной) мембраны”. Модель пригодна для объяснения широкого круга наблюдаемых явлений. При этом наблюдаемые явления описываются в рамках единого понятийного аппарата и не требуют специфических допущений для описания ...
... активность тиамина и некоторых его производных. За последние 20 лет наряду выяснением механизма основных реакций, в которых каталитическую роль играет ТДФ, стали накапливаться данные о высокой биологической активности других некоферментных производных тиамина. Отчетливо наметились два направления исследований: возможное, участие различных фосфорных эфиров витамина в активном переносе ...
... формами географической (территориально-механической) изоляции, известны и разные формы биологической изоляции, которые могут быть разбиты на три основные группы: эколого-этологическую, морфо-физиологическую и собственно генетическую. Биологическая изоляция приводит к уменьшению вероятности встречи особей разных полов в период размножения, снижению полового влечения и эффективности спаривания, к ...
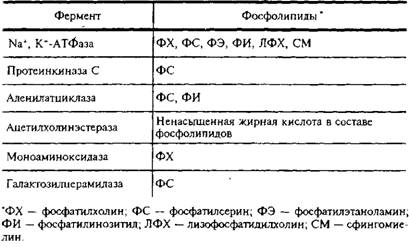
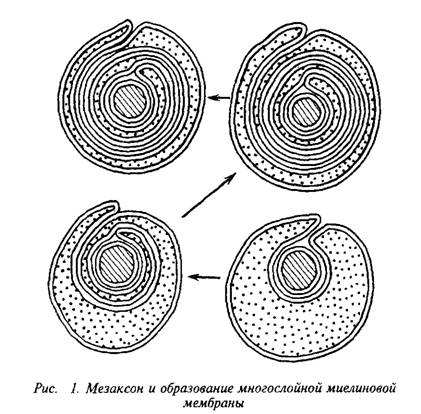
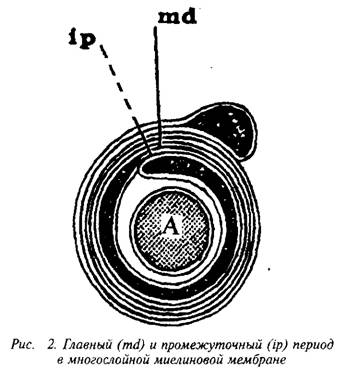
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
0 комментариев