Навигация
Особенности селекции люпина
121042
знака
14
таблиц
2
изображения
Введение
Узколистный люпин – ценная кормовая, сидеральная и пищевая культура.
В сложной ситуации сейчас находиться отечественное земледелие. Проводимые реформы в стране на многоукладность экономики и организации труда в сельском хозяйстве нарушили все его системы. Идет тенденция к снижению плодородия полей, все меньше применяются прогрессивные технологии возделывания сельскохозяйственных культур. В итоге – снижение урожайности и общий спад производства валовой продукции.
Для стабилизации растениеводческой продукции непременным условием является биологизация и экологизация земледелия на основе совершенствования структуры посевных площадей. Эта единственная возможность сохранить почвенное плодородие и прекратить обвальный спад производства продовольственного и фуражного зерна, грубых и сочных кормов.
Главным звеном биологической системы земледелия должно стать освоение севооборотов с короткой ротацией, предусматривающее увеличение бобовых культур в структуре посевов. Ведь для компенсации минеральных удобрений необходимы альтернативные источники питательных веществ, поскольку в таком земледелии уровень урожая формируют биологические факторы.
Говоря об активизации биологических факторов в системе земледелия путем посева одновидовых или смесей бобовых и злаковых растений, необходимо помнить, что биологизация должна означать не только замену антропогенных факторов естественными, но и представлять собой способ предотвращения возможных негативных последствий интенсификации растениеводства.
Таким образом, в существующем земледелии наличие бобовых трав в структуре посевов обязательно. Они – главный источник растительного белка и самые мощные азотфиксаторы. Трудно переоценить их пищевое, кормовое, агротехническое и экономическое значение. Одной из наиболее интересных таких культур является люпин узколистный.
Узколистный люпин имеет ряд объективных преимуществ:
Зерновая продуктивность (больше желтого люпина), высокая устойчивость к антракнозу и большая скороспелость (С.В. Бавровский, 1999). Достоинства узколистного люпина выдвигают его в ряд наиболее эффективных белоксинтезирующих растений. Это особенно важно для условий Нечерноземной зоны России, где возделывание других зернобобовых культур затруднено. В условиях экономического кризиса и ресурсного дефицита бобовые культуры имеют особое значение. Они – главный источник обогащения комовых рационов дешевым растительным белком (Т.Н. Слесарева, 1999). Производство растительного белка является центральной проблемой в решении продовольственных задач, т. к. продуктивность животноводства базируется, в основном, на растительном белке (В.С Лихачев, 1996).
Значение культуры люпина чрезвычайно велико не только как ценного кормового растения, но и как почвоулучшителя. Он накапливает в корнях и в надземной массе не менее 200 кг/га азота, обогащает активный слой почвы растворимыми формами фосфатов (В.И. Северов, 1996). Он обладает аллелопатическим и пролонгирующим действием. Люпин по праву можно назвать культурой многоцелевого использования. Его применяют в земледелии, животноводстве, лесоводстве, садоводстве, цветоводстве, почвозащитном деле, медицине, парфюмерии, лакокрасочной и пищевой промышленности. Экстракты из семян могут применяться при изготовлении медицинских препаратов, снижающих артериальное давление, регулирующих биоэнергетическую активность сердца, моторную и психическую активность. При современном уровне развития технологии переработки он является прекрасным сырьём для пищевого белка. Люпиновые белковые изоляты с успехом используются в хлебобулочной, макаронной, кондитерской, колбасной и мясоконсервной промышленности, в производстве диетических и лечебных профилактических продуктов.
Так белок люпина является хорошим компонентом для приготовления безглюциновых пищевых продуктов в виде печенья, пирожных и др. кондитерских изделий, обладающих диетическими и лечебно-профилактическими свойствами, включенных в питание школьников, студентов, спортсменов и космонавтов. Следовательно, узколистный люпин является культурой больших возможностей для многоцелевого использования: на удобрение и корм, на переработку для кормовых и пищевых целей. Наиболее полно потенциал люпина используется в Австралии, на втором мете – Беларусь. Дальше идут Польша, страны Африки, Испания, Чили, Россия и Германия. В Германии считают, что синий люпин (И.П. Такунов, 2001) должен спасти люпиносеяние в целом, называют его синим зонтиком надежды и «просветом для люпина». В Великобритании называют его «новым люпином» и «культурой будущего» и это действительно так. И у нас в России новые сорта узколистного люпина - это новый люпин.
В России люпиносеяние имело широкое развитие в шестидесятых годах прошлого века. Но в связи с массовым распространением болезней, низкими закупочными ценами на семена, малой урожайностью семян районированных тогда сортов, отличающихся продолжительным вегетационным периодом, площади под ним резко сократились. С появлением в настоящее время малоалколоидных сортов узколистного люпина наступил период его возрождения. Такой люпин является сравнительно новой культурой для С3 региона Росси и мало известен производственникам. Расширению его площадей препятствует хронический недостаток семян, так как возделываемые сорта не всегда созревают (Т.С. Козырев, 2001). Хотя устойчивое семеноводство узколистного люпина возможно в регионах с сумой активных температур свыше 1900 градусов по Цельсию за вегетационный период, пишет в журнале кормопроизводство №1 (2001) директор ВНИИ люпина, доктор сельскохозяйственных наук И.П. Такунов.
I. Обзор литературы. Хозяйственно-ценные признаки люпина узколистного
Основным достоинством люпина является его биологическая способность хорошо расти и развиваться на очень бедных песчаных почвах.
Обладая наивысшей азотфиксирующей способностью среди однолетних бобовых культур, люпин способен фиксировать при нормальном развитии в среднем 160-180 кг/га атмосферного азота. Исключительная нетребовательность люпина к почве объясняется также и тем, что его корневая система с помощью специальных выделений способна растворять труднорастворимые фосфорные соединения, недоступные для других культур, и накапливать в почве усвояемые формы фосфора. Кроме того, благодаря глубоко проникающей корневой системе, люпин усваивает в подпочвенных горизонтах промытые туда ранее и недоступные для других растений питательные вещества, в том числе калий и другие макро- и микроэлементы и, действуя как естественный насос, накапливает их в своей биомассе и вновь возвращает в пахотный слой почвы при запашке зеленой массы или пожнивных остатков и соломы. Люпин выступает здесь в роли биологического мелиоранта, предотвращая миграцию подвижных химических элементов в грунтовые воды, предохраняя тем самым от загрязнения окружающую среду. Зеленая масса люпина, запаханная в почву, эквивалентна по своему действию такому же количеству внесенного навоза, обогащает почву органическим веществом, повышает содержание в ней гумуса, значительно улучшает её физико-химические и биологические свойства и пищевой режим. Люпин оказывает большое влияние на улучшение качества сельскохозяйственной продукции. Запаханная зеленая масса люпина разлагается постепенно и в растениях не накапливается свободный азот, что мы часто наблюдаем при избыточном минеральном азотном питании растений. Кроме основной и неоценимой роли люпина в его внеконкурентной азотфиксирующей способности и возможности эффективного повышения плодородия бедных почв люпин обладает еще одним очень ценным свойством - является высокобелковым кормовым растением. Только соя по содержанию белка может сравниться с люпином. Белок люпина отличается высоким качеством, высокой переваримостью и из-за низкого содержания ингибиторов трипсина может использоваться в корм любым видам животных без предварительной термообработки, что обязательно при использовании в корм зерна сои.
Индекс МЕАА, используемый ФАО как показатель качества белка, составляет для люпина 70,65, в то время как МЕАА белка сои равен 69, 47.
Урожайность зеленой массы однолетних кормовых люпинов в зависимости от почвенно-климатических условий составляет в среднем 300-500 ц с 1 га, а в наиболее благоприятные годы достигает 700-800 и более центнеров. При урожайности зеленой массы люпина в 700 ц с гектара обеспечивается свыше 10 тонн сбора сухого вещества, в котором содержится более 2 тонн высококачественного кормового белка. Зеленая масса люпина хорошо поедается всеми видами животных как в свежем виде, так и в виде приготовленных кормов – силоса, сенажа, сена, витаминной муки, гранул, брикетов. Совместные посева люпина с ячменем для получения зернофуража и с овсом, кукурузой и другими культурами на зеленый корм и силос позволяют увеличить продуктивность пашни, повысить белковость получаемого корма и сбор белка с единицы площади и сбалансировать концентрированные и сочные корма по протеину непосредственно в поле без дополнительных затрат по их смешиванию при приготовлении кормов. Характерной особенностью белка люпина является полное отсутствие в нем проламинов, к которым относится глиадин и глютенин. В этой связи люпин является сырьем для создания безглютинных пищевых продуктов, обладающих диетическими и лечебно-профилактическими свойствами для детского питания.
Также люпин не вызывает аллергических реакций в организме, как это бывает при употреблении соевых продуктов.
Экстракты из семян горького люпина имеют потенциальную фармакологическую ценность и оказывают влияние на снижение артериального давления, биоэлектрическую активность сердца, моторную и психическую активность, не оказывая при этом наркотического действия. Алкалоид спартеин используется в качестве антиаритмичного средства. Возможно использование экстрактов люпина в качестве регуляторов роста.
Таким образом, люпин несет огромный биологический и экономический потенциал, который до настоящего времени полностью не используется.
Биологические особенности люпина
Однолетнее травянистое сильно облиственное растение. Стебель люпина узколистного прямостоячий. Ветвление начинается из пазух нижних листьев и идёт снизу вверх по главному стеблю. У позднеспелых форм преобладает длительное моноподиальное ветвление по всему растению снизу вверх, причем длина нижних побегов может достигать почти длины главного стебля. У скороспелых форм ветвление преимущественно симподиальное, верхушечное. Стебель не совсем выполненный, как бы гранёный, слабо опушённый, темно-зелёный, зелёный или светло-зелёный, иногда явно антоциановый – фиолетовый, фиолетово-коричневый, розовый.
Лист состоит из 7-8 узких, линейно-ланцетных листочков, согнутых по средней жилке – желобчатых. Соцветия небольшие, малоцветковые, с очередным расположением цветков. Венчик невзрачный, без запаха, различной окраски. Для всего вида L. Angustifolius характерна однородная окраска составных частей венчика – паруса, крыльев и лодочки.
Окраска венчика у узколистного люпина является одним из существенных морфологических признаков, которым, несомненно, необходимо пользоваться при подразделении этого вида на разновидности. Однако окраску венчика у L. Angustifolius никак нельзя рассматривать изолировано от окраски остальных частей растений (вегетативных органов и семян). Генетические исследования показывают, что у люпина узколистного часто наблюдается тесная коррелятивная зависимость окраски цветков, кожуры семян и вегетативных органов – стеблей и листьев.
Всё многообразие семян L. Angustifolius можно разделить по окраске семенной кожуры на 4 большие группы: белые, рыжие, коричневые, серые до черных. (Н.А. Майсурян, А.И. Атабекова.)
Морфогенез
В онтогенезе люпина можно выделить три основных периода: - первый: формирование и рост вегетативных органов (корней, стеблей и листьев), второй – формирование генеративных органов (соцветий и цветков) и третий период – формирование бобов и семян.
Видимые морфологические изменения отдельных органов и габитуса растений в процессе его развития принято отмечать по фенологическим фазам: появление всходов, розетка листьев, стеблевание, бутонизация, цветение и плодообразование, сизый боб, блестящий боб, спелый боб (Савичев, 1961; Наймарк, 1976)
Продолжительность фенофаз и этапов органогенеза зависит от генетических особенностей вида и сорта люпина, почвенно-климатических условий, технологических приемов возделывания и некоторых других факторов.
Рассмотрим этапы органогенеза у люпина по фенологическим фазам его развития.
Ø Прорастание семян
Для того, чтобы зародыш семени тронулся в рост, необходимо, чтобы под действием ферментов сложные запасные углеводистые и белковые соединения клеток семядолей стали доступными для его питания. Эти биохимические процессы протекают после поступления воды в семена и их набухания. Для прорастания семян требуется воды в 1,5-2 раза больше веса семян. При достаточно влажности почвы и температуре не ниже 10 градусов Цеьлсия семена набухают в течение 1-2 суток. Однако часть семян может приобретать свойство твердокаменности, которое препятствует поступлению воды в семена, и они могут длительное время оставаться в почве в непроросшем состоянии. Большинство авторов считают, что причиной твердокаменности семян является их пересушивание во время хранения, при котором происходит пересыхание пектиновых веществ палисадного слоя клеток эпидермиса семенной кожуры, причем этот процесс может быть обратимым и необратимым (Ржанова, 1970)
Для устранения твердокаменности семян и получения дружных всходов необходимо проводить скарификацию семян.
Момент прорастания семян принято считать за начало онтогенеза растения, он соответствует 1 этапу органогенеза. Характеризуется формированием конуса нарастания и первых зародышевых листьев почки.
Ø Фаза листовой розетки
Фаза листовой розетки наступает рослее появления всходов с момента образования двух настоящих свернувшихся листочков, укрывающих точку роста стебля, соответствует II этапу органогенеза. У основание конуса нарастания формируются габитус растения в целом.
Фаза прикорневой розетки у люпина узколистного практически отсутствует. После появления всходов и выноса семядолей на поверхность почвы начинается активный рост стебля параллельно с ростом корневой системы. В этом плане узколистный люпин по темпам первоначального роста выгодно отличается от желтого и белого люпинов и является наиболее скороспелым видом.
Ø Фаза стеблевания
В разные годы фаза стеблевания длится от 9 до 23 дней и соответствует III-IV-V этапам органогенеза.
У узколистного люпина процессы формирования и роста побега осуществляются синхронно. (Борисова, 1990)
На четвертом этапе органогенеза происходит формирование генеративной сферы растения. У основания верхушечного конуса нарастания в пазухах листовых зачатков закладываются конусы нарастания ветвей второго порядка. На пятом этапе органогенеза формируются органы цветка.
Ø Фаза бутонизации
Данная фаза характеризуется появлением бутонов на центральной кисти и длится до начала цветения в течение 8-15 дней и соответствует VI-VII-VIII этапам органогенеза.
На шестом этапе органогенеза происходит микро- и мегаспорогенез. На седьмом этапе идет формирование мужского и женского гаметофитов (двухядерной пыльцы и зародышевого мешка семяпочки) На восьмом этапе органогенеза происходит процесс опыления.
Ø Фаза цветения и плодообразования
Данная фаза наступает с начала раскрытия первого цветка на первой мутовке главной кисти и заканчивается образованием бобов на центральной кисти, длится 9-13 дней и соответствует IX этапу органогенеза. На этом этапе происходит опыление и оплодотворение только у перекрестноопыляющихся видов люпина. У самоопыляющихся видов цветение происходит после того, как произошло самоопыление в нераскрытом бутоне, и сводится лишь к раскрытию цветка, биологическое значение которого пока еще мало изучено (Ржанова, 1970)
Период цветения люпина может затягиваться за счет цветения боковых ветвей.
Ø Фаза сизого боба или зернообразования
наступает после цветения при образовании темно-зеленых бобиков, длиной 2-4 сантиметра, густо покрытых волосками, придающими бобам сизовато-серебристый оттенок. В эту фазу рост главного стебля в высоту приостанавливается. Дальнейший прирост урожая зеленой массы идет за счет роста боковых ветвей и налива зерна. Фаза сизого боба длится от 16 до 23 дней и соответствует X этапу органогенеза, который характеризуется начальной дифференциацией зародыша семени, образованием проэмбрио и закладкой семядолей и конуса нарастания почечки семени. К концу X этапа происходит полный налив зерна и бобы приобретают свойственную им форму (Савичев, 1961)
Ø Фаза блестящего бобы или зеленой спелости
Соответствует XI этапу органогенеза. Зерно приобретает надлежащую форму. В этот период идёт усиленная аттракция питательных веществ из створок боба в семена. Темно-зеленая окраска бобов меняется на зеленую и светло-зеленую, бобы начинают блестеть. На одиннадцатом этапе органогенеза происходит формирование зародышевых листьев в почечке. В этой фазе заканчивается прирост биомассы и наиболее целесообразно убирать люпин на силос и запахивать на зеленое удобрение. К концу этой фазы при необходимости можно применять дефолиацию для ускорения уборки посевов на семенные цели.
Ø Фаза спелого боба
Соответствует XII этапу органогенеза и связана с созреванием семян. В это время происходит окончательный отток питательных веществ в семядоли из всех органов растения, в том числе из створок боба и семенной кожуры. В семядолях протекают биохимические процессы по превращению простых органических соединений в сложные запасные вещества (Ржанова, 1970) В период восковой спелости зерна окраска семядолей белесая, корешок зародыша желтеет. В дальнейшем семядоли также приобретают желтый цвет. Бобы становятся плоскими с ярко очерченными местами расположения семян. Створки бобов вначале буреют, затем приобретают желто-коричневый цвет. В сухую погоду растения люпина полностью засыхают и наступает пора уборки. В дождливую погоду часть листьев на боковых ветвях может быть еще зеленой, что осложняет уборку и требует дополнительных затрат для проведения дефолиации.
В.С. Шевелуха (1992) отмечает в онтогенезе люпина два основных периода роста: 1) медленный рост надземных и быстрый рост подземных органов в фазу розетки листьев; 2) быстрый рост стебля и корневой системы в фазы стеблевания и бутонизации. Второй период является критическим в отношении влаги.
Значение и особенности развития корневой системы
В системе целого растения корневую систему можно рассматривать не только в качестве «энергетического блока», отводящего ассимиляты листа и, таким образом, регулирующего фотосинтез, но и учитывая факт обеспечения растения через корни различными минеральными и физиологически активными веществами, - органа, определяющего слаженность и интенсивность течения всего комплекса процессов метаболизма. К настоящему времени накоплен большой фактический материал, указывающий на то, что фотосинтетическая активность листьев в большой мере зависит от функционального состояния корневой системы растения.
Учитывая, что корневая система люпина не только поглощает минеральные питательные вещества и воду из почвенного раствора, но и синтезирует в корневых клубеньках самый дефицитный из элементов питания – азот, беря его из неорганического сырьевого источника – воздух, особенности развития корневой системы люпина представляют особый интерес. Рост корней в глубину у всех возделываемых однолетних видов люпина происходит за счет стержневого корня, который достигает глубины 1,5-2 и более метров. Наиболее интенсивный рост корней и побега происходит в фазу стеблевния и бутонизации. В фазу цветения рост главного стебля прекращается, а рост корней несколько замедляется, но продолжается почти до полного созревания семян (Филимонова, 1962; Борисова, 1990) Интенсивный рост боковых корней первого прядка начинается в фазе розетки со времени появления третьего настоящего листа, а массовое образование боковых корней второго порядка – с фазы 7-8 настоящих листьев (Духанин, 1973).
Как известно, люпин, по сравнению с другими сельскохозяйственными культурами, слабо отзывается не только на основные элементы минерального питания, но и на некоторые микроэлементы. Причина этого явления кроется в высокой усвояющей способности корней люпина по отношению к труднодоступным элементам питания почвы (Д.И. Прянишников, 1965)
Усваивающую способность корней растений, в частности, могут характеризовать показатели удельной поглощающей поверхности и ёмкости катионного обмена. Растения, у которых эти показатели выше, в большей степени способны поглощать труднодоступные элементы минерального питания из почвы.
Образование клубеньков на корнях люпина начинается с фазы 3-4 пар настоящих листьев. Их число и размеры постепенно возрастают и достигают максимума в фазу цветения. Более 70% клубеньков располагается на главном стержневом корне и около 30% - на боковых корнях первого порядка.
И.П. Такунов (1996)
Отношение люпина к внешним условиям
- Требования к теплу
Центром происхождения возделываемых однолетних видов люпина являются страны Средиземноморского побережья, поэтому по своей природе люпин относится к числу сравнительно теплолюбивых растений, хотя каждый вид имеет свои определенные особенности. Узколистный люпин является менее требовательным. Оптимальная температура прорастания семян узколистного люпина является +9…+12ºС (минимальная +2…+4ºС).
К.И. Савичев (1961) отмечает, что семена люпина, положенные в непрогретую влажную почву, начинают прорастать при температуре 1-2 градуса, однако всходы в этом случае появляются только на 20-26 день, а при посеве в хорошо прогретую почву всходят на 5-6 день. Температура почвы и воздуха влияет как на продолжительность периода посев-всходы, так и на полевую всхожесть семян люпина. При низкой температуре почвы прорастание семян затягивается, семена подвергаются воздействию различных почвенных патогенов, некоторые семена плесневеют и теряют всхожесть. По данным В.Н. Степанова, для прорастания семян и появления всходов у узколистного люпина требуется сума температур 110ºС.
В.Н. Дюбин (1981), обобщив данные Госсорсети и материалы станций Госкомгидромета, пришел к выводу, что сумма средних суточных температур, необходимая для появления всходов, от даты посева составляют для узколистного люпина - 150ºС. Чем выше среднесуточная температура воздуха (при прочих оптимальных условиях), тем быстрее появляются всходы у всех видов люпина. Люпин наиболее чувствителен к весенним заморозкам в фазу семядольных листочков, но понижение температуры до -2..-3ºС выносят все виды люпина. В фазе 4-6 настоящих листьев всходы люпина узколистного выдерживают понижение температуры до -7…-9ºС. ( Бадина, 1974) К.И Савичев (1961) сообщает, что при температуре 4-10 градусов у узколистного люпина происходит стадия яровизации. Повышение температуры после прохождения стадии яровизации благоприятно сказывается на их дальнейшем развитии. Чем выше температура при наличии влаги в почве, тем быстрее идет рост и развитие люпина. Понижение температуры замедляет и удлиняет все фазы развития, а при температуре ниже 10 градусов прохождение фаз развития у люпина как бы приостанавливается. Большое значение для люпина имеет температурный режим в межфазный период цветение-созревание семян, на который приходится 42-50% суммы температур всего вегетационного периода. Чем выше средняя температура воздуха в этот период, тем быстрее созревает люпин. С понижением температуры при продвижении люпина с юга на север продолжительность этого периода увеличивается и созревание семян затягивается (Бадина, 1974) Температура и влажность почвы оказывает наибольшее влияние на формирование урожая семян и вегетативной массы люпина. Более высокая температура при достаточной увлажненности почвы в период стеблевание-цветение способствует большему накоплению зеленой массы и образованию бобов на растении.
Следует отметить, что у люпина, посеянного семенами, созревание которых протекало при пониженных температурах, вегетационный период бывает значительно короче, чем у растений посевов, произведённых семенами, созревшими при более высоких температурах. Это позволяет предполагать, что при созревании семян в условиях пониженных температур стадия яровизации их проходит на корню материнского растения.
- Требования к свету
Все однолетние зернобобовые культуры светолюбивы, и люпин узколистный в том числе. Недостаток света вызывает усиленный рост стеблей, вытягивание их, слабое развитие корневой системы, плохое цветение и плодоношение. В урожае уменьшается содержание белка, так как снижение освещенности влияет на активность фотосинтеза, в связи с чем ощущается недостаток углеводов для биосинтеза белков.
Гелиотропизм – свойство поворачивать свои листья перпендикулярно падающим солнечным лучам в течение всего светового дня, свойственно люпину. Благодаря большой площади листовой поверхности – до 100 га на 1 га растения люпина имеют наибольший коэффициент использования солнечной энергии – 4,79, в то время как у пшеницы он равен 2,68, у ржи – 2,42, у клевера – 2,18, у вики – 1,98 (Бадина, 1974) В зависимости от стадии вегетации потребность в освещенности бывает различной. Так в молодом возрасте люпин лучше переносит затемнение, чем в более поздние периоды жизни (Орлов, 1986) Наиболее чувствителен к свету люпин в период формирования бобов и созревания семян. Наряду с интенсивностью освещения большое влияние на развитие растений оказывает его продолжительность или соотношение длины дня и ночи. Е.К. Алексеев (1986) отмечает, что выделить фактор фотопериодической реакции люпина в «чистом виде» трудно, потому что влияние её переплетается с действием других факторов. Наблюдения над произрастанием люпина узколистного в нашей стране дают основание считать их растениями длинного дня.
Регулировать освещенность люпиновых ценозов можно с помощью технологических приёмов: изменяя сроки и способы посева, густоту стояния растений, подбирая такие компоненты для смешанных посевов, которые в наименьшей степени затеняли бы люпин.
- Требования к влаге
Культурные виды люпина – влаголюбивые растения. В засушливых районах они не растут. Это объясняется тем, что они образуют большую вегетативную массу. Транспирационный коэффициент люпина равен 600-700. Однако мощная корневая система люпина, проникая глубоко в почву и подпочву, даёт возможность растениям добывать воду из глубины и тем самым меньше зависеть от условий атмосферного увлажнения; но это когда растения уже хорошо разовьются, а в начальный период они очень требовательны к наличию влаги. Указывают два критических периода чувствительности люпина к недостатку воды: период прорастания семян и период формирования на растениях генеративных органов, особенно с фазы бутонизации – весь период цветения до образования блестящих бобов (Алексеев, 1968) Часто семена люпина трудно набухают. Трудность набухания объясняется наличием в их кожуре твердого палисадного слоя клеток, препятствующего всасыванию воды, и химическим составом самой оболочки. Это нежелательное свойство, снижающее качество посевного материала, называется твердокаменностью (Циклаури, 1962) При большом количестве твердокаменных семян вполне здоровые, свежие семена дают очень низкий процент всхожести. В очень ярко выраженной форме проявляется недостаток влаги в период бутонизация - цветение у малоалколоидных сортов узколистного люпина, сопровождаемый настолько сильным опадением цветков, что почти полностью отсутствует урожай семян. Наиболее благоприятной для узколистного люпина влажность 60-75% от полной влагоемкости почвы. В то же время переувлажнение почвы также неблагоприятно сказывается на развитии растений, как и недостаток воды: вегетационный период удлиняется люпин может совсем не дать семян. Избыток влаги для люпина также неблагоприятен. В годы с избыточным увлажнением период вегетации люпина удлиняется, созревание семян задерживается, усиливается поражение растений грибными болезнями.
Повышение влажности почвы во время бутонизации и цветения приводит к увеличению урожайности семян и зеленой массы, не задерживая при этом созревания. Положительное влияние оказывают и непродолжительные дожди, выпадающие в период созревания семян, - они обуславливают повышение крупности зерна и белковости семян (Алексеев, 1986) При повышении влажности почвы не только увеличивается урожайность, но и снижается в растениях количество алкалоидов.
- Требования к почве
Люпины способны произрастать на самых бедных почвах, однако заметно повышают урожаи при всяком улучшении почвенного плодородия.
Узколистный люпин предпочитает более связанные почвы от супесчаных до среднесуглинистых (на песках растёт плохо).
Невысокая требовательность люпина к почвенным условиям слагается из нескольких его особенностей. Это прежде всего наличие мощной корневой системы стержневого типа с боковыми ответвлениями. Способность люпина фиксировать атмосферный азот при помощи специфических рас клубеньковых бактерий, присущих только ему. Являясь энергичным азотонакопителем, люпин независим от запасов азота в почве, что также выступает фактором слабой требовательности к почвенному плодородию.
Повышенные требования люпина к доступу кислорода к корням и аэрации почвы связаны с тем, что клубеньковые бактерии образуются в основном на корнях, расположенных в поверхностных слоях почвы, а по мере углубления корневой системы их количество резко снижается.
- Отношение к кальцию
Как и любой другой элемент кальций необходим для питания люпина и отсутствие его в почвенном растворе вызывает физиологические расстройства.
«Кальциефобность» люпина проявляется в задержке поступления железа в растения и заболевании их хлорозом.(Алексеев, 1968)
Особое положение люпина среди других бобовых растений по отношению к извести выражается в том, что он очень интенсивно поглощает кальций, особенно в период от всходов до начала цветения.
В растениях люпина кальций накапливается главным образом в стеблях или листьях, в то время как в семенах и створках бобов содержание его наименьшее. Согласно А. Уоллесу (1966), поглощение кальция бобовыми происходит, по-видимому, против градиента концентрации и в отличие от поглощения калия не зависит от температуры, pH и других условий.
На бедных фосфором почвах при отсутствии фосфорных удобрений или даже при совместном внесении извести и фосфоритной муки одной из главной причин отрицательного действия извести на люпин является фосфорное голодание растений. Это обуславливается переходом фосфора в малорастворимое состояние под влиянием внесенной извести и неспособностью люпина в ранний период его вегетации использовать фосфор труднорастворимых фосфатов. Поэтому устранение отрицательного действия извести на люпин путём внесения в почву бора или путём замены части CaCO3 на MgCO3 возможно лишь при достаточной обеспеченности этой культуры фосфором или при внесении в качестве фосфорного удобрения суперфосфата. Основываясь на современных данных, можно констатировать, что избыток кальция действует на люпин угнетающе. Чтобы избежать вредного действия извести, лучше проводить известкование непосредственно под люпин, используя медленность взаимодействия извести с почвой.
- Требования к удобрениям
Люпин относится к культурам, не предъявляющим особых требований к условиям питания в силу особенностей своей корневой системы. А наличие корневых клубеньков означает то, что люпин не зависит от запасов азота почвы.
Отношение к фосфору
Корневая система люпина обладает способностью хорошо усваивать фосфор труднорастворимых фосфатов не только аморфного, но и кристаллического строения. Кроме того, воздействие люпина на фосфорит объясняется растворяющим действием его корневой системы, что сопровождается повышением концентрации водородных ионов окружающей среде.
Таким образом, люпин хорошо растёт и развивается на источниках фосфорного питания любой доступности.
Отношение к калию
По отношению к калию люпин является наиболее требовательным из всех бобовых растений. Он энергично поглощает калий и при низком, и при высоком его содержании в почве, особенно при выращивании на рыхлопесчаных почвах.
Калий повышает оводненность тканей и способствует экономному использованию воды, благодаря чему повышается устойчивость люпина к недостатку почвенной влаги. Также способствует усилению углеводного обмена в растениях и повышению синтеза белковых соединений. В зависимости от почвенной разности калийные удобрения повышают содержание белка в семенах люпина на 4-5%, валовый сбор его в урожае – на 2-3 ц/га. Калий снижает алкалоидность люпина, особенно в засушливые годы.
Из микроэлементов люпин чаще всего реагирует на недостаток молибдена и бора, которые участвуют в белковом синтезе.
Молибден играет определяющую роль в ассимиляции азота.
Бор наряду с калием увеличивает оводненность тканей и интенсивность фотосинтеза.
Болезни люпина
Почти всегда в посевах люпина можно встретить растения, пораженные болезнями. Наиболее вредоносны для люпина грибковые болезни и вирусы и в меньшей степени бактериозы.
Фузариоз
Заболевание это вызывается паразитными грибками из рода фузариум. В начале у корневой шейки люпина появляется темно-коричневый налет, а затем слизь. Слизь, проникая в ткани растения, вызывает загнивание корней, потемнение стебля, пожелтение и покраснение листьев, их увядание и опадение. Пораженные растения отстают в росте, преждевременно подсыхают, часть бобов у них осыпается, а развившиеся бобики дают щуплое зерно. При сильном поражении растения могут погибнуть еще до образования плодов. (К.И. Саввичев)
Исследованиями установлено, что основным источником фузариозной инфекции являются почва и растительные остатки, в связи с чем частое или бессменное возделывание люпина на полях, высокое насыщение севооборотов с короткой ротацией (2-3 поля люпина в 4-5 польных севооборотах) приводит к накоплению инфекции в почве. (И. П. Такунов, 1996). Потери урожая от фузариоза могут оставлять: зеленой массы до 45% и семян - до 58%.
Таблица №1. Доноры и источники устойчивости люпина узколистного к фузариозу
N каталога ВИР (VIR) Название сорта, образца Происхождение Тип устойчивости Группа созревания
Люпин узколистный (L. angustifolius)
2831 Лаф-рбс/2 Белоруссия У 3
2832 Лаф-рбс/3 -//- У 3
2833 Лаф-рбс/1 -//- У 3
2834 Лаф-рбс/9 -//- У 3
2840 Лаф-рбс/10 -//- У 3
2686 Вада-14 -//- У 4
2750 БСХА-892 -//- У 4
2748 БСХА-640 -//- У 4
2682 Апва-27 -//- У 4
2703 ДС-397 -//- У 4
2841 ДМ-75/84 -//- У 3
2877 Линия Н 56-23 Россия УУ 3
2878 Линия Н 58-10 -//- УУ 3
2673 Elvas Португалия У 4
Группы созревания: 1 — крайне ранний (менее 71 дня); 2 — очень ранний; 3 — полуранний ; 4 — ранний ; 5 — среднеспелый; 6 — среднепоздний.
В последние годы в Белоруссии созданы новые сорта узколистного люпина Арва, Вада и К-2750, которые показали высокую устойчивость к фузариозу во всех регионах испытания (Белоруссия, Киев, Новосибирск и С.-Петербург). Устойчивость узколистного люпина к фузариозу контролируется двумя доминантными генами. При создании новых фузариозоустойчивых сортов узколистного люпина автором используется два различных подхода. При подходе 1 два восприимчивых родителя скрещивали между собой и устойчивые формы отбирали в гибридных потомствах, у которых наблюдалась комплементарность взаимодействия неаллельных генов устойчивости. Большинство сортов узколистного люпина в Белоруссии было создано при использовании подхода 1: Данко, Арва, Вала, Силена, Миртан, Першацвет.
При подходе 2 в качестве доноров устойчивости использовали дикие формы, в частности, образец Apendrion из Греции, в скрещиваниях с культурными сортами с последующим отбором фузариозоустойчивых растений. В итоге были созданы устойчивые сорта и линии Вика-65, Ксерап-653, DG-45 и др. (Чекалин Н.М).
Антракноз является очень вредоносной болезнью в Средиземноморских странах, Андах, Австралии, особенно в регионах с влажным климатом для всех возделываемых видов люпина. В последние годы, вероятно, в связи с глобальным потеплением климата возбудитель антракноза стал представлять реальную угрозу возделыванию люпина в люпиносеющих регионах Украины, России и Белоруссии. Болезнь поражает листья, черешки, стебли, бобы и семена, образуя темные, впалые повреждения с розово-оранжевой споруляцией в центре. Споры распространяются, главным образом, с инфицированными семенами. Болезнь впервые была идентифицирована в США в 1939 г. на узколистном люпине. Несколько лет спустя среди дикой гермплазмы из Португалии были найдены устойчивые линии и эта устойчивость была интродуцирована в возделываемые сорта узколистного люпина. (Чекалин Н.М).
Антракноз оказывает резкое отрицательное влияние на родуктивность культуры: у растений отмечается заметное отставание в росте, уменьшение количества бобов, снижение урожайности зеленой массы и зерна, ухудшние посевных качеств (И. П. Такунов, 1996).
В 90-е годы антракноз становится главным препятствием в производстве люпина в Европе, Южной Америке, Австралии и Новой Зеландии. Распространению болезни способствовало образование новых патотипов, лучше приспособленных к засушливым условиям. Только несколько линий отмечены, как более устойчивые по сравнению с обычными сортами после новой вспышки антракноза.
Так, при инокуляции сорта белого люпина Rio Maior 14 изолятами возбудителя антракноза, а 35 линий люпина белого и 32 линий люпина узколистного — изолятом 96А4 была отмечена следующая картина: все изоляты поражали белый люпин от низкого до очень высокого уровня; линии белого люпина поражались все — балл поражения 1,1—4,5; линии узколистного люпина очень сильно различались по степени поражения — от 0 до 5 баллов. Высокоустойчивой к антракнозу оказалась линия узколистного люпина Alt 97—16, относительно устойчивыми были линии белого люпина Prima Baer, SAL 101, Bunheiro-Murtosa и Jambrina 1.
Sweetingham et al. указывает, что доминантный ген устойчивости к антракнозу был идентифицирован у люпина узколистного в Австралии. У сорта Rancher устойчивость контролируется доминантным геном An. Обнаружено 2 расы болезни VCG-1 и VCG-2, которые встречаются в Европе, причем высоковирулентная раса VCG-2 была распространена по всем континентам с семенами в 70-ые гг. XX столетия. (Чекалин Н.М).
Важной биологической особенностью патогенна является сравнительно узкая специализация, высокая приспособленность к растению-хозяину, способность поражать все органы растения с образованием в пораженной ткани колоссального репродуктивного спороношения (до 1 млн конидий на 1 мм ткани), развитие возбудителя с максимальной скоростью и появление эпифитотий (Цветкова, 1995)
Исходя из того, что основной путь распространения антракноза – зараженные семена, важнейшим звеном в системе защиты от этого заболевания является обеззараживания семян (И.П. Такунов, 1996).
Серая листовая пятнистость (Stemphylium botriosum Wallr., St. solani Weber) — аббр. СЛП. Впервые обнаружена на юго-востоке США в 50-е гг. Симптомы — небольшие округлые пятна на листьях и иногда на стеблях и бобах. У восприимчивых сортов узколистного люпина происходит полная дефолиация. Устойчивые образцы этого вида люпина найдены среди диких форм в Португалии и мутантных селекционных линий из шт. Джорджия, США (87). Гены устойчивости к СЛП были интродуцированы в австралийские сорта узколистного люпина Marri (1975) и Illyarrie (1979). Благодаря этим сортам, эпидемия болезни стала редким явлением в Западной Австралии, в силу их устойчивости и раннего созревания. (Чекалин Н.М).
Цератофороз или бурая пятнистость листьев
Широко распространенное грибковое заболевание люпина. Поражаются листья, стебли, бобы и семена всех культивируемых видов люпина. Первые признаки появляются в мае в виде точечных черно-бурых пятен. Со временем пятна увеличиваются и сливаются, листья преждевременно засыхают и опадают. При сильном развитии болезни растения сплошь бурые, как бы обожженные. Вредоностность цератофороза заключается в преждевременном опадении пораженных листьев, значительном снижении урожая зеленой массы и семян из-за нарушения работы фотосинтетического аппарата. При сильном поражении примерно 25% растений не образует плоды. Основным источником первичной инфекции служат пораженные цератофорозом растительные остатки и зараженная почва, где гриб длительное время сохраняет свою жизнеспособность. Поэтому применение агротехнических приемов, ограничивающих накопление патогенна в почве – севооборот, глубокая зяблевая вспашка, уничтожение послеубоорчных растительных остатков, - способствует значительному снижению развития болезни на посевах (И. П. Такунов, 1996).
Мучнистая роса является наиболее распространенной болезнью листьев люпина. Обычно появляется во второй половине лета, на пораженных листьях образуется белый налет; при сильном развитии болезни налетом покрываются не только листья, но и стебли люпина. Сильнее болезнь развивается в засушливые годы и на сухих открытых местах.
Источником инфекции служат многолетний люпин и пораженные растительные остатки. Из агротехнических мер борьбы применяются ранние сроки сева, удаление пораженных растительных остатков. Хорошие результаты дает обработка посевов растворами фунгицидов.
Болезни корней у люпина вызываются возбудителями различных видов: выпревание (черная ножка) — грибами родов Pythium spp., Fusarium spp., Rhizoctonia spp. Черная ножка вызывает гибель проростков до или после всходов из-за инфекции коры или гипокотиля. У R. solani идентифицировано 3 штамма в Австралии — AG 11, AG 2-2 и AG 2-1, вызывающие гниль гипокотилей у узколистного люпина. Встречается довольно редко. (Чекалин Н.М).
Вирусные болезни люпина
Наиболее часто встречаются два вида вирусных заболеваний люпина: вирусное израстание или узколистности и вирусное побурение. Возбудителем узколистности является вирус желтой мозаики фасоли (ВЖМФ), побурение – вирус огуречной мозаики (ВОМ-1).
На узколистном люпине вирусное израстание начинается с листьев, они становятся сначала розовыми, а потом пурпурными. Верхушка соцветия крючкообразно изгибается и полностью увядает. Растение погибает.
Побурение, вызываемое вирусом огуречной мозаики, особенно резко проявляется на люпине в фазе цветения. При этом наблюдается резкое укорачивание междоузлий и интенсивный рост боковых побегов. На стебле и во влагалищах листьев появляются бурые некротические полосы, верхушка растения привядает, а центральный цветонос крючкообразно изгибается. Стебель и черешки становятся хрупкими и растение вскоре погибает. Оно имеет обугленный вид и внешне похоже на погибшее от фузариоза. Ущерб, причиняемый заболеванием, в целом незначительный.
Задачи и методы селекции
В настоящее время наиболее перспективным направлением в селекционной работе является комплексный подход, основанный на применении знаний частной генетики как качественных, так и количественных признаков, использовании в работе созданных или выделенных генетических доноров хозяйственно ценных признаков и моделей статической генетики и биометрии.
Это позволяет решать следующие задачи:
Ø Изучать селекционную ценность отдельных генов, групп и созданных на их основе доноров с последующим использованием лучших из них в селекции.
Ø Создавать трансгрессивные формы за счет вовлечения в селекционный процесс образцов, имеющих аллельные различия в контроле образцов. Трансгрессивные формы можно эффективно выявлять на основе разработанного в отделе зерновых бобовых культур ВИР эколого-географического подхода.
Ø Создавать и выявлять исходный материал, обладающий групповой устойчивостью к разным видам, штаммам и расам патогенов за счет аккумуляции в создаваемых генотипах разных генов устойчивости, путем испытания исходного материала в контрастных условиях среды и подбором пар для скрещивания на основе эколого-географического подхода.
Ø Изучать качественный состав популяции возбудителей болезней в конкретных условиях, а также степень их вирулентности, что позволит целенаправленно создавать устойчивые сорта для каждого региона и при необходимости своевременно проводить сортосмену.
Ø Целенаправленно получать новые мутанты с применением скрининга и рекомбинации новых сложных ассоциаций генов.
Ценное свойство люпина – его способность благодаря симбиозу с клубеньковыми бактериями фиксировать атмосферный азот. За счет этого повышают плодородие почвы биологическим путем, экономят дорогостоящие минеральные удобрения, Выращивают экологически чистую продукцию и не загрязняют окружающую среду. В связи с изложенным селекция на высокую азотфиксирующую способность очень перспективна.
Основными направлениями селекции люпина на повышение симбиотической азотфиксации являются:
Ø Подбор эффективных селекционных штаммов клубеньковых бактерий к уже созданным высокопродуктивным сортам люпина;
Ø Создание высокопродуктивных сортов с высокой клубнеобразующей способностью, активнотью и эффективностью симбиотической азотфиксации при взаимодействии с соответствующими штаммами клубеньковых бактерий в условиях естественного низкого содержания или отсутствия в почве минерального азота;
Ø Поиск комплементарных симбиозов, способных выбирать из почвы определенный и эффективный штамм бактерий;
Ø Создание сортов, избегающих конкуренции со спонтанной микрофлорой (без применения препаратов);
Ø Выведении сортов, образующих большое количество клубеньков с высокой активностью и эффективностью биологической азотфиксации при наличии в почве достаточного количества минерального азота, но не использующих его;
Ø Нахождение образцов, которые на минеральном фоне обеспечивают положительный эффект при применении ризоторфина;
Ø Создание эффективных комплементарных симбиозов макро- и микросимбионтов для разных селекционных задач.
Исключительно актуальная в настоящее время проблема селекции в отношении всех видов люпина – создание сортов, устойчивых к вирусным и грибным болезням, особенно фузариозу и антракнозу. Неудачи последних лет, приведшие к сокращению посевных площадей под люпином, связаны с этими заболеваниями.
Важной задачей для создания сортов однолетних видов является селекция на скороспелость и стабильную семенную продуктивность. Скороспелые сорта должны обладать рядом других положительных качеств. К ним прежде всего следует отнести быстрые темпы роста люпина, что позволит упростить борьбу с сорной растительностью. Признак нерастрескиваемости бобов должен быть придан всем новым сортам люпина узколистного. Кроме того, новые сорта этого вида должны обладать симподиальным типом ветвления.
Новое перспективное направление – создание форм и сортов с детерминантным типом роста стебля и детерминированным ветвлением.
Основные направления в селекции люпина узколистного следующие:
Создание скороспелых сортов интенсивного типа, с высокой и стабильной урожайностью семян до 2,5-4,0 и зеленой массы 35,0-50,0 т/га, с высоким содержанием белка в семенах (до 35%) и зеленой массе (17-20%), с нерастрескивающимися бобами, устойчивых к грибным и вирусным заболеваниям, к заморозкам до -10 градусов Цельсия и кратковременной засухе.
Методы селекции
Методы отбора
В течение многих веков человек использовал естественные популяции люпина, имеющие всегда большое разнообразие по морфологическим, биологическим и хозяйственным признакам. На основе таких популяций под влиянием сначала естественного, а потом и искусственного отбора, народная селекция в странах Средиземноморья и на Американсом континенте создавала высокопродуктивные местные сорта разных видов люпина. Многие сорта создаются аналитическим путем с помощью индивидуального и индивидуально-семейственного отборов и в настоящее время. Путем непосредственного отбора из районированных сортов и коллекционных образцов, особенно гибридного происхождения, были созданы многие известные в мире сорта (Академический 1, Быстрорастущий 4 и др,). Большое значение в селекции люпина при работе с популяциями имеет многократный отбор. Он позволяет в короткий срок разложить популяцию на отдельные более однородные группы, между лучшими формами которых наиболее эффективно проводить в дальнейшем скрещивания (Таранухо, 1980).
При селекции на высокую азотфиксирующую способность наиболее информативной является оценка большого набора образцов различного эколого-географического происхождения с последующим отбором на разных фонах: без внесения минерального азота; без минерального азота, но с инокуляцией семян перспективными штаммами клубеньковых бактерий; на минеральном азоте; а также на минеральном азоте с инокуляцией семян Rhizobium lupini. Такие исследования с одним и тем же набором образцов желательно провести в разных регионах. В результате открывается перспектива выявить диапазон и закономерности изменчивости показателей симбиотической азотфиксации, а также отобрать ценный исходный материал для селекции (Картузова и др., 1991).
Разные формы отборов являются неотъемлемым элементом на всех этапах селекционного процесса.
Внутривидовая гибридизация
В настоящее время основные методы селекции - экспериментальная и спонтанная внутривидовая гибридизация. Чаще всего используют ступенчатое скрещивание, которое позволяет объединить в одном генотипе ценные признаки и свойства, присущие многим исходным формам. При проведении гибридизации и работе с гибридным материалом применяют бекроссы и рекурентные схемы селекции, диаллельные, ступенчатые и возвратные скрещивания. Классическим примером высокоэффективного применения ступенчатой гибридизации, при которой поэтапно к уже созданным формам дополняются недостающие признаки других сортов, является создание серии немецких сортов Вайко (Майсурян, Атабекова, 1974; Таранухо, 1979). С целью передачи новым сортам отдельных ценных признаков без отрицательных свойств донора применяют бекроссы и рекурентные схемы селекции. Однако отбор используют и в процессе гибридизации. С использованием метода педигри уже в третьнм поколении получают новые константные формы, возникающие в результате проявления комбинационной изменчивости. В последние годы получает распространение метод отбора одного семени в потомстве ( метод SSD).
В целях избежания восстановления синтеза алкалоидов при перекрестном опылении, разработаны способы отбора и селекции перекрестноопыляющихся видов на стабильную безалкалоидность. Способ отбора форм люпина многолетнего на безалкалоидность (Чекалин, Курлович, 1989) включает выделение безалкалоидных растений, их размножение, отличающееся тем, что с целью повышения эффективности процесса селекции за счет получения популяции, стабильной по признаку безалкалоидности, после выделения безалкалоидных растений между ними осуществляют парные скрещивания. Все потомство, полученное от каждого скрещивания, выращивают изолированно и среди него отбирают в качестве исходного материала популяцию, в которой полностью отсутствуют алкалоидные растения, и ее размножают изолированно.
Использование предлагаемого способа позволило создать доноры люпина многолистного с достаточно высокой стабильностью признака безалкалоидности (ВИР-1, ВИР-3, ВИР-4, ВИР-5, ВИР-29). Недостатком способа является трудоемкость. так как для выделения популяции стабильной по признаку безалкалоидности необходимо привлекать в контролируемые парные скрещивания большое количество безалкалоидных растений. К тому же, значительная их часть используется неэффективно, поскольку растения оказываются несовместимыми по признаку безалкалоидности и в процессе работы выбраковываются. Безалкалоидные растения, привлекаемые в скрещивания, как правило, обладают рядом отрицательных признаков и свойств (низкая продуктивность, неустойчивость к неблагоприятным факторам среды, болезням, вредителям), что не позволяет получить высококачественный селекционный материал.
Упрощение приведенного способа и получение высокопродуктивных безалкалоидных форм (Курлович, Чекалин, 1992) достигается тем, что каждое выделенное безалкалоидное растение скрещивают с алкалоидной формой, обладающей ценными признаками, а безалкалоидные нерасщепляющиеся популяции выделяют во втором гибридном поколении для дальнейшего их использования в селекционном процессе. С использованием способа создан высокопродуктивный донор безалкалоидности ВИР-41, а также переданный в государственное сортоиспытание первый сорт кормового многолистного люпина Первенец.
Предложенные способы исключают возможность выщепления алкалоидных растений в безалкалоидных популяциях за счет комплементарного взаимодействия неаллельных генов, обуславливающих безалкалоидность. Однако в посевах кормовых сортов различных видов люпина может иметь место частичное выщепление горьких растений за счет обратных мутаций, а также в результате переопыления безалкалоидных форм с дикорастущими на близком расстоянии алкалоидными растениями. В связи с этим во всех звеньях семеноводства безалкалоидных сортов необходимо строго соблюдать пространственную изоляцию между разными образцами, а также осуществлять постоянный контроль уровня алкалоидности. Выявленные горькие растения необходимо в обязательном порядке удалять до цвете.
Изложенные способы могут быть эффективно использованы в се- лекционной работе со многими перекрестноопыляемыми видами люпина: Lupinus polyphyllus Lindl., L. perennis L., L.arboreus Sims., L.mutabilis Sweet., L.succulentus Dougl., L.elegans H.B.K. и др.
При селекции на высокую урожайность, устойчивость, качество и по другим сложным количественным признакам эффективны подходы В.А. Драгавцева (1993) по компьютерному отбору двух (или нескольких) лучших для гибридизации родителей. Методами идентификации нужных генетических систем в F2 в камере или в поле (при той же типичной динамике лимитирующих факторов) отбираются уникальные растения, несущие в себе генетические системы обоих родителей, являющиеся трансгрессиями и родоначальниками ценных сортов.
Отдаленная гибридизация
Полиморфный род Lupinus L. объединяет две самостоятельные, обособленные друг от друга группы видов восточного и западного полушарий. Различное их эволюционное развитие привело к возникновению между ними изолирующих барьеров в отношении скрещиваемости. Люпины из этих двух групп межвидовых гибридов не образуют. Также остаются бесплодными попытки скрестить между собой большинство видов люпина с восточного полушария с различным числом хромосом. Однако многие виды люпина с западного полушария с 2n=48 достаточно легко скрещиваются между собой и дают фертильное или достаточно фертильное потомство. Кроме того, многие из признанных ныне видов люпина западного полушария являются скрытыми гибридами или полугибридами (Майсурян, Атабекова, 1974). В литературе наиболее полно описаны гибриды: L.ornatus x L.mutabilis, L.pubescens x L.Hartwegii, L.mutabilis x L.elegans, L.mutabilis x L. albococcineus, L.nootkatensis x L.arboreus, L.arboreus x L.hartwegii, L.mutabilis x L.douglasii (Kazimierski, 1960, 1963; Kazimierski, Nowacki, 1961а, Kazimierski, Kazimierska, 1970; Kazimierska, 1970; Майсурян, Атабекова, 1974).
Что касается видов люпина из западного полушария (subgen. Lupinus), то к настоящему времени удалось скрестить между собой только люпин желтый (Lupinus luteus) с обоими подвидами люпина испанского ssp.hispanicus и ssp.bicolor (Lamberts, 1955, 1958; Kazimierski, Kazimierska, 1970; Майсурян, Атабекова, 1974; Swiecicki, 1988). Более жизнеспособное и ценное в селекционном отношении потомство получается от скрещивания L.luteus x L.hispanicus ssp.bicolor (синоним - L.rothmaleri), а также L.luteus с гибридами ssp.hispanicus ssp.bicolor (Cortero, Arrieta, 1988).
Кроме того, в литературе как межвидовые описаны гибриды люпина узколистного с льнолистным и позднецветущим, а также люпина белого с люпином Вавилова и югославским (Kazimierski, 1960; Майсурян, Атабекова, 1974). Однако, как выяснилось позднее (Gladstones, 1970, 1974; Курлович, Станкевич, 1990), такие гибриды не являются межвидовыми, поскольку люпины льнолистный и позднецветущий представляют собой всего лишь отдельные экотипы люпина узколистного, а люпин Вавилова и Югославский - один из подвидов люпина белого (subsp.graecus).
Эмбриологические исследования показали (Kazimierski, 1961; Kazimierska, 1970), что низкая фертильность некоторых межвидовых гибридов является следствием отклонений в мейозе и различных аномалий в процессе развития зародышевого мешка. Однако полученные положительные результаты несомненно представляют теоретический интерес и указывают на родственные связи и степень близости между различными видами люпина. Достижения современной и будущей биотехнологии несомненно помогут преодолеть барьеры нескрещиваемости между многими видами люпина, что откроет новые возможности и перспективы культуры (Sator, 1984).
Мутагенез
Большинство сортов люпина создано с использованием спонтанных или индуцированных мутантов. Первые кормовые сорта желтого, узколистног и белого люпина были получены в Германии в 1927-1928 гг. в результате размножения выявленных мутантных безалкалоидных растений (Sengbusch, 1931, 1942; Майсурян, Атабекова, 1974; Коновалов и др., 1990). В дальнейшем, естественные мутанты использовались в гибридизации для улучшения многих показателей (белая окраска и быстрая набухаемость семян, нерастрескиваемость и неопушенность бобов, быстрые темпы начального роста). В результате, путем ступенчатой гибридизации, была создана серия знаменитых немецких сортов люпина желтого под названием Вайко.
В последние годы для индуцирования мутаций широко используют химические мутагены: этиленимин, диэтилсульфат, нитрозометилмочевина, гидроксиламин и др., а также различные виды ионизирующих излучений. Метод искусственного мутагенеза у люпина перспективен тем, что позволяет значительно расширить спектр и увеличить частоту появления форм с измененной наследственной основой у этого сравнительно молодого культурного растения. Больше полезных мутаций отмечают при воздействии химическими мутагенами в сравнении физическими факторами (Солодюк, 1971). Их концентрации и экспозиции зависят от объекта (сортовых особенностей, сухие или набухшие семена, кормовые или горькие формы, разные виды люпина).
В наших исследованиях наиболее эффективным химическим мутагеном оказалась нитрозометилмочевины (НММ) в концентрации 0,8 мМоль. Сначала семена замачивали в воде в течение 8 часов, а затем в течение 3 часов содержали в растворе нитрозометилмочевины.
В опытах Г.А. Дебелого и А.В. Зекунова (1977) также установлена высокая мутагенная эффективность ННМ в сравнении с гамма-лучами и ЭИ. Наибольшая частота и широкий спектр (8 типов) мутации получены в концентрации НММ 0,015% у сорта Н-846. На сорте Немчиновский синий выделились по мутагенной эффективности две концентрации - 0,015 (12,5% мутантов, 11 типов мутаций) и 0,014 (10,1% мутантов и 12 типов мутаций). Изучавшиеся на двух сортах три концентрации ЭИ (0,01; 0,02; 0,034) оказались близкими по мутагенной эффективности.
Между контрастными сортами люпина наблюдались специфические различия. Так, у алкалоидного сорта Немчиновский синий был значительно шире спектр мутаций, для него были характерны рецессивные мутации по окраске цветка и семян, реже встречались мутации по размеру бобов и семян. У малоалкалоидного сорта Н-846 наиболее широкий спектр мутаций наблюдался по признакам, определяющим габитус и структуру стебля (карлики, компактные, штамбовые и др.). В равной мере выделены у обоих сортов мутанты с бобами, устойчивыми к растрескиванию и лучшей выполненостью. Позднеспелые мутанты выделены у обоих сортов, а скороспелые - лишь у сорта Немчиновский синий. У обоих сортов обнаружены мутации с измененным ритмом роста (мутанты быстрорастущие и медленнорастущие по сравнению с исходным сортом). У алкалоидного сорта выделены малоалкалоидные мутанты. У обоих сортов на естественном и искусственных инфекционных фонах отмечены мутанты, устойчивые к фузариозу.
По J.Hackbarth (1957b), мутанты подразделяются на восемь групп: 1) листового аппарата - эта группа включает хлорофильные мутанты, мутанты с изменением интенсивности окраски, антоциановой пигментации и размера листьев; 2) цветка - с измененной окраской цветка; 3) семян - с измененной формой, размером и окраской семян; 4) боба - с измененным размером боба (крупнобобовые, мелкобобовые), выполненностью, с различной степенью устойчивости к растрескиванию; 5) физиологические - позднеспелые и скороспелые, с измененным ритмом роста; 6) биохимические - с изменением по содержанию белка и алкалоидов; 7) устойчивости - к болезням и другим стрессовым факторам; 8) всего растения - мутанты высокорослые, карлики, и с измененным габитусом (компактные, без ветвей, штамбовые).
При использовании физических факторов (рентгеновского и гамма-излучения в дозах от 150 до 250 Гр) учитывают мощность дозы и биологические особенности признаков, по которым желательно найти мутантную форму (Коновалов и др., 1990). Измененные наследственно формы можно отбирать в течение ряда поколений, начиная с М2.
В результате использования искусственного мутагенеза получены селекционно-ценные формы люпина с детерминированным ветвлением, фасциированным стеблем, нерастрескивающимися бобами, низким содержанием алкалоидов, повышенным содержанием белка и масла, отличающиеся скороспелостью и термонейтральностью. Они явились исходным материалом для создания кормовых сортов люпина белого - Киевский мутант, Старт, Горизонт, Пищевой, Приморский, Днепр, Олешка; люпина узколистного - Ладный, Дикаф 14; люпина желтого - Житомирский, Юбилейный, Искорость и др.
Полиплоидия
Этот метод, дающий положительные результаты в селекции многих сельскохозяйственных культур, в практической селекционной работе с люпином пока не используется в связи со сложностями, возникающими из-за большого числа хромосом в клетках этой культуры. Однако в Польше oпубликована работа (Kazimierska, Kazimierski, 1969) по изучению биологических особенностей гаплоидов люпина желтого, которые были выявлены среди химерных диплоидных растений. Они, а также полученные из них дигаплоиды оказались менее жизнеспособными в сравнении с диплоидными формами, имели меньшую массу семян и продуктивность. В последующих исследованиях (Kazimierski, Kazimierska, 1992) выявлены гаплоидные и тетраплоидные формы у люпина белого. Наибольший теоретический и практический интерес представляют тетраплоиды (2n=4х=100). Хотя они имели в бобе меньшее количество семян в сравнении с диплоидными формами, масса семян у них оказалась в два и более раза выше. В результате по урожайности они не уступали обычным диплоидным формам. Меньшее количество семян в бобе у тетраплоидов авторы объясняют нарушениями в процессе мейоза. Приведенные эколого-географическое изучение и отборы позволили выделить наиболее продуктивные растения, которые предполагается использовать в селекции.
Естественные тетраплоиды иногда встречаются также среди дикорастущих люпинов из западного полушария - Lupinus perennis L. (Майсурян, Атабекова, 1974).
Оценка селекционного материала на инфекционных фонах и путем искусственного заражения
Люпин подвержен многочисленным заболеваниям, среди которых самые распространенные и вредоносные - фузариоз и антракноз. Болезни вызывают возбудители, относящиеся к разным видам родов Fusarium Lk. и Colletotrichum gloesporioides Penz. Их соотношение и состав неодинаковы в различных почвенно-климатических зонах. Сорта люпина, устойчивые к фузариозу и антракнозу в одних регионах, часто оказываются восприимчивыми в других. Оценка селекционных образцов люпина на устойчивость к фузариозу проводится на инфекционных фонах.
Методика создания инфекционных фонов к почвенным патогенам (фузариозы) основана на накоплении инфекции в почве (Гешеле, 1964; Корнейчук, 1985). Для испытания люпина приемлемы самые распространенные методы - двух-трехлетняя монокультура сильновосприимчивого сорта, внесение в почву измельченных частей пораженных растений, а также метод искусственного внесения в почву размноженных на питательной среде патогенов (Корнейчук, 1985). В качестве питательного субстрата могут служить зерна хлебных злаков, особенно овса и проса. Можно использовать и зерна других культур и их смеси. Реже применяют внесение в почву чистой культуры патогена, приготовленной на жидкой питательной среде (Овчинникова, Андрюхина, 1984).
В зависимости от конкретных задач, возможны разные подходы к изучению устойчивости люпина к фузариозу. Например, мировая коллекция люпинов ВИР, включающая более 50 видов как средиземноморского, так и американского происхождения, в 1971-1989 гг. получила первичную оценку по устойчивости на комплексном инфекционном фоне Новозыбковского филиала ВИУА (Киселев и др., 1988). Этот фон создавали, внося в почву измельченную массу больных фузариозом растений, собранных в разных зонах люпиносеяния, а также чистую культуру разных видов и штаммов возбудителя. На таком фоне возможно первоначальное испытание всех видов люпина.
Однако, для более углубленного изучения фузариозоустойчивости образцов каждого вида люпина, целесообразно создание специального фона со специфичным для этого вида формами возбудителя. Так, в Украинском НИИ земледелия разработана методика создания инфекционного фона для оценки люпина желтого к фузариозному увяданию на основе выделенных в условиях Украины высоковирулентных штаммов (82, 66, 89, 90, 213) Fusarium oxysporum Schlecht. (Корнейчук, 1985). В НПО "Подмосковье" и НПО зернобобовых и крупяных культур с целью создания такого фона для люпина узколистного и белого выделяют изоляты с больных фузариозом растений, определяют их видовой состав и патогенность. Для закладки фона используют наиболее вирулентные из них (Дебелый и др., 1983; Ларионова, Овчинникова, 1987).
Важным направлением борьбы с фузариозом может быть создание сортов, обладающих групповой устойчивостью к разным видам и штаммам патогена. Как показали исследования (Курлович и др., 1990), для этого эффективно испытывать один и тот же исходный материал для селекции в контрастных условиях среды на разных фонах, уже созданных во многих селекционных учреждениях страны. Таким путем удалось выявить материал, устойчивый к фузариозу в разных регионах, а также изучить вариабильность признака устойчивости в зависимости от конкретных условий.
Значительные различия в поражении большинства сортов люпина в разных регионах люпиносеяния и на разных фонах обусловливают необходимость изучения качественного состава популяции возбудителя болезни в конкретных условиях, а также степень ее вирулентности. Обладая такой информацией, располагая данными об устойчивости материала к тому или иному составу патогенов, в перспективе будет возможно целенаправленно создавать устойчивые сорта для каждого региона, а при необходимости своевременно проводить сортосмену.
Организация разных инфекционных фузариозных фонов и методы оценки материала изложены в работах А.В. Хотяновича, Г.С. Муромцева (1975), Н.С. Корнейчука (1985), Л.И. Ларионовой, Б.С. Курловича (1990).
Что касается антракноза, то инфекция этого патогена передается в основном через семена и сохраняется в течение зимы на пораженных растительных остатках. Болезнь вызывает на листьях своеобразные пятна, которые на стеблях и бобах переходят в язвы. В местах поражения после дождя может выступать слизистая масса конидий. С брызгами дождя инокулюм распространяется и инфицирует соседние здоровые растения. В связи с этим, методика создания инфекционного фона к этому заболеванию несколько иная. Обычно тестируемые растения обрызгивают раствором, в котором содержатся в большом количестве наиболее вирулентные для данного региона штаммы патогена. Поскольку так же как и у фузариоза расовый и штаммовый состав возбудителей антракноза в разных местах не одинаков, поиск устойчивого материала следует вести в том самом месте, где ведется селекция культуры, используя для заражения наиболее вирулентные в местных условиях расы и штаммы. Подтверждением этому являются результаты испытания образцов люпина белого и желтого, привезенных нами из Бразилии, где имели место сильные эпифитотии этого заболевания, и на их фоне в Бразилии выделены устойчивые формы (Курлович и др., 1989). Однако их испытание в разных эколого-географических условиях стран СНГ при заражении местными штаммами и расами патогена в большинстве случаев не выявило у них высокой устойчивости к заболеванию.
Вирусная узколистность известна во всех районах люпиносеяния. На основании изучения биологических, морфологических, физиологических и других свойств вирусных изолятов в условиях, например, Украины (Курлович и др., 1989) выделены два штамма наиболее распространенного вируса желтой мозаики фасоли (ВЖМФ), различающиеся по степени вирулентности (слабо- и сильновирулентный). Также имело место распространение смешанной инфекции ВЖМФ и вируса огуречной мозаики (Cucumis virus 1, Smith.), но не столь распространенной, как инфекция, вызванная только ВЖМФ (Phaseolus virus 2, Smith.). В связи с этим изучение образцов на пораженность узколистностью и поиск источников обычно проводится в два этапа: на фоне естественного заражения (первичная оценка) и на фоне искусственного заражения ВЖМФ (основная оценка).
(http://lupin-rus.blogspot.com/2006/07/lupin-breeding.html)
В настоящее время наиболее перспективным направлением в селекционной работе является комплексный подход, основанный на применении знаний частной генетики как качественных так и количественных признаков, использовании в работе созданных или выделенных генетических доноров хозяйственно ценных признаков и моделей статической генетики и биометрии.
Это позволяет решать следующие задачи:
Ø Изучать селекционную ценность отдельных генов, групп и созданных на их основе доноров с последующим использованием лучших из них в селекции.
Ø Создавать трансгрессивные формы за счет вовлечения в селекционный процесс образцов, имеющих аллельные различия в контроле образцов. Трансгрессивные формы можно эффективно выявлять на основе разработанного в отделе зерновых бобовых культур ВИР эколого-географического подхода.
Ø Создавать и выявлять исходный материал, обладающий групповой устойчивостью к разным видам, штаммам и расам патогенов за счет аккумуляции в создаваемых генотипах разных генов устойчивости, путем испытания исходного материала в контрастных условиях среды и подбором пар для скрещивания на основе эколого-географического подхода.
Ø Изучать качественный состав популяции возбудителей болезней в конкретных условиях, а также степень их вирулентности, что позволит целенаправленно создавать устойчивые сорта для каждого региона и при необходимости своевременно проводить сортосмену.
Ø Целенаправленно получать новые мутанты с применением скрининга и рекомбинации новых сложных ассоциаций генов.
Ценное свойство люпина – его способность благодаря симбиозу с клубеньковыми бактериями фиксировать атмосферный азот. За счет этого повышают плодородие почвы биологическим путем, экономят дорогостоящие минеральные удобрения, Выращивают экологически чистую продукцию и не загрязняют окружающую среду. В связи с изложенным селекция на высокую азотфиксирующую способность очень перспективна.
Основными направлениями селекции люпина на повышение симбиотической азотфиксации являются:
Ø Подбор эффективных селекционных штаммов клубеньковых бактерий к уже созданным высокопродуктивным сортам люпина;
Ø Создание высокопродуктивных сортов с высокой клубнеобразующей способностью, активнотью и эффективностью симбиотической азотфиксации при взаимодействии с соответствующими штаммами клубеньковых бактерий в условиях естественного низкого содержания или отсутствия в почве минерального азота;
Ø Поиск комплементарных симбиозов, способных выбирать из почвы определенный и эффективный штамм бактерий;
Ø Создание сортов, избегающих конкуренции со спонтанной микрофлорой (без применения препаратов);
Ø Выведении сортов, образующих большое количество клубеньков с высокой активностью и эффективностью биологической азотфиксации при наличии в почве достаточного количества минерального азота, но не использующих его;
Ø Нахождение образцов, которые на минеральном фоне обеспечивают положительный эффект при применении ризоторфина;
Ø Создание эффективных комплементарных симбиозов макро- и микросимбионтов для разных селекционных задач.
Исключительно актуальная в настоящее время проблема селекции в отношении всех видов люпина – создание сортов, устойчивых к вирусным и грибным болезням, особенно фузариозу и антракнозу. Неудачи последних лет, приведшие к сокращению посевных площадей под люпином, связаны с этими заболеваниями.
Важной задачей для создания сортов однолетних видов является селекция на скороспелость и стабильную семенную продуктивность. Скороспелые сорта должны обладать рядом других положительных качеств. К ним прежде всего следует отнести быстрые темпы роста люпина, что позволит упростить борьбу с сорной растительностью. Признак нерастрескиваемости бобов должен быть придан всем новым сортам люпина узколистного. Кроме того, новые сорта этого вида должны обладать симподиальным типом ветвления.
Новое перспективное направление – создание форм и сортов с детерминантным типом роста стебля и детерминированным ветвлением.
Основные направления в селекции люпина узколистного следующие:
Создание скороспелых сортов интенсивного типа, с высокой и стабильной урожайностью семян до 2,5-4,0 и зеленой массы 35,0-50,0 т/га, с высоким содержанием белка в семенах (до 35%) и зеленой массе (17-20%), с нерастрескивающимися бобами, устойчивых к грибным и вирусным заболеваниям, к заморозкам до -10 градусов Цельсия и кратковременной засухе. (Курлович Б.С., ВИР, 1995г.)
Ценный исходный материал
В диком состоянии в настоящее время люпин произрастает в странах по всему периметру средиземного моря и в северо-восточной части Африки. (Купцов Н. С., Такунов И. П. 2006).
Наиболее распространенными и изученными, представляющими перспективу как исходный материал при селекции, являются следующие виды: L. Luteus L. 2n=52 (Желтый); L. Rothmaleri Klinr. 2n=52 (Ротмалара); L. albus 2n=50 (Белый); L. vavilovi Atab Et. Maiss. 2n=50 (Вавилова); L. Angostifolius L. 2n=40 (Узколистный); L. linifolius Roth 2n=40 (Льнолистный); L. Opsianthus Atab. Et Maiss. 2n=40 (Поздноцветущий); L. Pilosus L. 2n=40 (Волосистый); L. digitatus Forsk. 2n=40 (Пальчатый); L. hirsutus L. 2n=24 (Мохнатый); L. mutabilis Sweet. 2n=48 (Изменчивый) и др.
При селекции любого культурного растения весьма важно использовать подходящий исходный материал. От этого зависит значительная доля успеха в селекционной работе.
Несмотря на то, что селекция узколистного люпина очень молода (начало было положено в конце сороковых годов), её результативность значительна. Селекционным путём достигнуты скороспелость, неполегаемость, высокая и стабильная продуктивность (урожай семян и вегетативной массы), малоалкалоидность и другие хозяйственно ценные признаки культуры. Для продвижения северных границ ареала узколистного люпина необходимы более скороспелые сорта – с продолжительностью периода вегетации 85..95 дней. Создание таких сортов возможно на основе измененного морфотипа растений – с генетическим блокированием бокового ветвления. Исследованиями НИИСХ ЦР НЗ установлено, что формы с ограниченным ветвлением центрального побега (эпигональное) или боковых побегов I порядка (детерминантные) имеют несомненное преимущество по скороспелости и семенной продуктивности в зонах с коротким вегетационным периодом и невысокой (менее 1900ºС) суммой положительных температур.
Впервые такой принципиально новый морфобиологический тип растений люпина узколистного описан Г.А. Дебелым и А.В. Зекуновым. Курлович изучал характер наследования детерминантности ветвления и установил, что у одних образцов этот признак контролируется рецессивными аллелями двух генов (deb 1, deb 2), у других доминантными аллелями двух других генов (Deb 3 и Deb 4). При этом во всех случаях происходит неаллельное взаимодействие генов, на характер которого оказывает модифицирующее влияние окружающая среда. Установлено, что гибридизация детерминантов с формами обычного ветвления позволяет получить ценные рекомбинаты для создания ультраскороспелых сортов зернофуражного использования. Селекция узколистного люпина в данном направлении успешно развивается в Австралии и Белоруссии. В России созданы и в Государственный реестр селекционных достижений два детерминантных сорта Ладный и Дикаф 14 .
В ранних комбинациях скрещиваний чаще использовали сорта Москвского селекцентра Ладный, Дикаф 14 и сортообразец селекции Белорусского НИИ земледелия и кормов Ланадекс 1.
Сорт Ладный – индуцированный мутант ценного в селекционном отношении сорта Немчиновский 846, обладающего высокой комбинационной способностью. Детерминация его ветвления контролируется рецессивными генами deb 1, deb 2.
Детерминированный с нерастрескивающимися бобами сорт Дикаф 14 получен селекционерами ТСХА и НИИСХ ЦР НЗ от скрещивания Фрост с Ланадекс 1. Материнская форма этого сорта хорошо изучена. Сорт Фрост имеет несколько ценных генов, An и gl контролируют соответственно устойчивость к антракнозу и к серой листовой пятнистости, ius – малоалкалоидность, leus – белые цветки и семена, s – мягкокожурность семян. Ланадекс 1 – это принципиально новая форма растения, имеющая в пазухах листьев вместо боковых побегов цветки. Этот призак контролируется доминантными аллелями Deb 3 и Deb 4.
По многообразию рекомбинантов выделился гибрид Брянский 35 Х Дикаф 12. В первом поколении гибрид отличался высокой зерновой продуктивностью и дружным созреванием, что свойственно рекомбинантам. Во втором поколении он выщепил продуктивные по семенам формы с разной степенью детерминации ветвления, растение с пигментированными бобами, что свидетельствует о наличии гена нерастрескиваемости 1е. Очень многообразен он по высоте растений. Гибрид гетерозиготен по алкалоидности. Для последующей селекционной работы из него отобраны интересные формы с комплексом хозяйственно полезных признаков.
В селекционных питомниках по зерновой продуктивности выделился колосовидный селекционный номер (СН). Брянский 1272 комбинации ДПСЛXДанко. По количеству семян он превзошёл стандартный сорт Дикаф 14 на 54%, а по массе – на 146%. По этим показателям он не уступает высокопродуктивному сорту обычного морфотипа Брянский 123. Эта константная форма имеет довольно развитые нижние побеги, которые отходят от стебля под углом, близким 90º и служат своеобразными подпорками для центрального стебля. По длине вегетационного периода Брянский 1272 относится к ультраскороспелой группе. Период вегетации варьирует по годам от 75 до 80 дней. Отличительная особенность СН Брянский 1272 – устойчивость к растрескиванию бобов. Она сопровождается проявлением маркерного признака этого свойства – розовая окраска створок бобов. К сожалению сорт не стабилен по концентрации алкалоидов. Требуется дальнейшая селекционная доработка по этому признаку.
Одна из константных форм обратной комбинации (ДанкоXДПСЛ) СН Брянский 1289 имеет блокировку ветвления на уровне побегов первого порядка. По семенной продуктивности новая форма превзошла стандарт на 80%. Она отличается дружным созреванием и мелкосемянностью: масса 1000 семян равна 111г.
По скороспелости в селекционных питомниках выделили группу номеров колосовидного морфотипа комбинации Ладный Х Детер синий: СН 297, СН 298. По зерновой продуктивности они незначительно превосходят стандартный сорт обычного морфотипа Брянский 123, но созревают почти на месяц раньше его.
При оценке на устойчивость к фузариозу форм с детерминированным типом ветвления выделяются линии гибрида Брянский 35 Х ДII розовый. В течение трех лет их устойчивость на жестком инфекционном фоне бла стабильной и колебалась в пределах 67,7…71,8% при 26,8% у сорта-индикатора Узколистного 109.
В некоторых скрещиваниях использовали мутантную форму ТСХА – 5М2-263. Она имеет фасциированный утолщенный стебель, сильно утолщенный цветонос с большим количеством расположенных на нем цветков. Эта форма привносит в гибрид повышенную зерновую продуктивность, хотя обладает и рядом отрицательных свойств: алкалоидностью, неустойчивостью к полеганию и грибным болезням.
Значительный селекционный интерес представляют гибридные комбинации Детерминант 1 х 5М2-263. Так линия 932 в пятом поколении константна по морфологическим признакам. Характеризуется сиреневой окраской цветков, белыми мелкими семенами. Лучшие растения формируют до 100 бобов.
С 1994 г. В скрещивания вовлечены новые сортообразцы селекции Белорусского НИИ земледелия и кормов – Метель и Першацвет, которые кроме детерминации ветвления обладают устойчивостью к осыпанию семян, контролируемой тремя рецессивными генами.
Сорт Метель - типичный представитель метельчатого морфотипа. Его генеративная сфера похожа на прямостоячую метелку злаков. Бобы расположены на растении эпигонально и компактно.
Анализ гибридов первого поколения выявил их высокую семенную продуктивность. Так, у лучших из них, в том числе Брянский Л-3 х Метель, Метель х Брянский 35, количество семян одного растения варьировало от 113 до 140, а их масса превышала 20 г. В F1 доминирует скороспелость. Гибриды созрели одновременно и дружно, в основном за 90…95 дней. В то же время родительские формы созревали крайне неравномерно как в пределах сорта, так и отдельно взятого растения.
Очень ранним и дружным созреванием отличался гибрид Першацвет х СН 118/94. Материнская форма сорт Першацвет – типичный представитель колосовидного морфотипа. Его генеративная сфера напоминает колос, состоящий из бобов. Отцовская форма СН 118/94 – высокорослый детер, более позднеспелый из-за растянутого периода всходы-цветение. В гибриде доминирует скороспелость материнской формы.
В заключение можно констатировать наличие обширного фонда исходного и селекционного материала для создания новых высокопродуктивных сортов люпина узколистного. (П.А. Агеева, 1996)
Исходным материалом для создания новых сортов люпина узколистного могут служить местные сорта в районах их длительного использования, селекционные сорта и мутантные формы, дикорастущие формы, произрастающие в генцентрах, так из диких видов селекции на повышение продуктивности представляют интерес люпины льнолистный, мохнатый, поздноцветущий, дающие плодовитые гибриды при скрещивании их с люпином узколистным (Купцов Н.С., Такунов И.П., 2006).
II. Экспериментальная часть
Похожие работы

... масса 1000 зерен была выше у сорта Радужный, как в 2005 году – 165 г., так и в 2008 году – 159 г. Все изучаемые сорта люпина узколистного показали довольно высокую зерновую продуктивность в условиях северной лесостепи Челябинской области (таблица 9). Сорт Белозерный, показал хорошие результаты по урожайности в 2005 году, но в 2008 г. при менее благоприятных метеоусловиях, он оказался не устойчив ...
... роста, сокращению завязываемости плодов и понижению урожая в 3-4 раза, по сравнению с урожаем в оптимальных условиях. В период налива и созревания семян потребность во влаге снижается. Люпин – светолюбивое растение. Для него характерно явление гелиотропизма – листья всегда повернуты перпендикулярно лучам Солнца. Размещение более 100 растений на 1м² способствует быстрому отмиранию нижних ...
... на устойчивость к стрессовым факторам среды (холод, инфекция) с помощью биохимических методов (изменчивость изоферментов пероксидазы, содержание сахаров, крахмала, фенольных соединений). Исследования по селекции и сортоизучению яблони проводятся сотрудниками З.А. Козловской, Г.М. Марудо, Ю.Г. Марчук, Е.Н. Бирюк, Т.А. Гашенко, С.И. Ярмоличем и В.М. Евдокименко. Телефон: +375-17-506-66-38. Груша: В ...
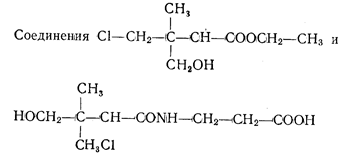
... фазы развития. Кроме того, применение указанных гаметоцидов задерживало прохождение фенологических фаз у растений, а также выколашивание части стеблей. Однако, несмотря на отрицательные побочные действия гаметоцидов, применение их в ближайшей перспективе может открыть путь к реализации эффекта гетерозиса у зерновых (особенно пшеницы), технических, овощных и кормовых культур. Поэтому весьма ...
0 комментариев