Формальная модель нейрона и нейросети
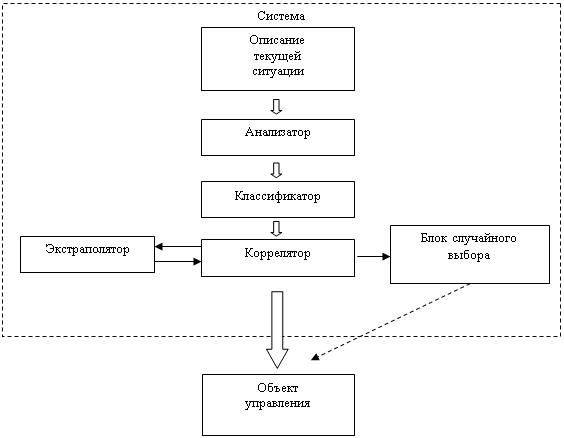
Краткое описание метода автономного адаптивного управления
Основные понятия и обозначения
Моделирование среды
Аппарат формирования и распознавания образов
Задача построения ФРО
Распознавание пространственно-временных образов
Система построения и исследования нейронных сетей СПИНС
Конструкторы сетей. Библиотеки шаблонов
Организация вычислений в сети
Реализация модели среды
Навигация
Аппарат формирования и распознавания образов
Нейросетевая реализация системы
61847
знаков
3
таблицы
10
изображений
3. Аппарат формирования и распознавания образов.
3.1. Биологический нейрон.
На рис. 3.1.1, взятом из [Turchin] представлен в упрощенном виде биологический нейрон. Схематично его можно разделить на три части: тело клетки, содержащее ядро и клеточную протоплазму; дендриты – древовидные отростки, служащие входами нейрона; аксон, или нервное волокно, - единственный выход нейрона, представляющий собой длинный цилиндрический отросток, ветвящийся на конце. Для описания формальной модели нейрона выделим следующие факты:
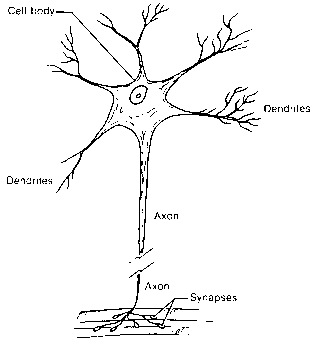
Рис. 3.1.1
В любой момент возможны лишь два состояния волокна: наличие импульса и его отсутствие, так называемый закон “все или ничего”.
Передача выходного сигнала с аксона предыдущего нейрона на дендриты или прямо на тело следующего нейрона осуществляется в специальных образованиях – синапсах. Входные сигналы суммируются с синаптическими задержками и в зависимости от суммарного потенциала генерируется либо нет выходной импульс – спайк.
3.2. Формальная модель нейрона.
Впервые формальная логическая модель нейрона была введена Маккалоком и Питтсом в 1948 году [Маккалок] и с тех пор было предложено огромное количество моделей. Но все они предназначены для решения в основном задач распознавания и классификации образов. Можно указать целый ряд основных отличий предлагаемой в данной работе модели и уже существующих. Во-первых, в классических моделях всегда присутствует “учитель” или “супервизор”, подстраивающий параметры сети по определенному алгоритму, предлагаемый же нейрон должен подстраиваться “сам” в зависимости от “увиденной” им последовательности входных векторов. Формально говоря, при работе нейрона должна использоваться только информация с его входов. Во-вторых, в предложенной модели нет вещественных весов и взвешенной суммации по этим весам, что является большим плюсом при создании нейрочипа и модельных вычислениях, поскольку целочисленная арифметика выполняется всегда быстрее, чем рациональная и проще в реализации. Главное же отличие предлагаемой модели состоит в цели применения. C помощью нее решаются все задачи управляющей системы: формирование и распознавание образов (ФРО), распознавание и запоминание закономерностей (БЗ), анализ информации БЗ и выбор действий (БПР), в отличии от классических моделей, где решается только первая задача.
Важной задачей ФРО для автономных систем также является не только распознавание образов, но и их хранение или запоминание. Это следует из автономности системы, т.к. для неавтономных систем распознанные образы могут храниться и использоваться вне системы. Вообще говоря, проблему запоминания можно решить множеством способов. Например, один из известных способов – организовать кольцо из нейронов, в котором сигнал мог бы прецессировать до бесконечности или в течении некоторого времени в случае затухания. В последнем случае система приобретает новое полезное свойство “забывчивости”, которое, как известно, присуще биологическим системам и позволяет более рационально и экономно использовать ресурсы, т.к. ненужная или малоиспользуемая информация просто “забывается”. Эксперименты проводились с формальной моделью без памяти, но очевидно, что она нужна. Нами предлагается ввести так называемую синаптическую память, т.е. способность запоминать входной сигнал в синаптическом блоке.
В данной работе мы используем нейрон из [Жданов2], который модифицирован в соответствии с [Братищев]. Мы приведем лишь краткое описание. На рис. 3.2.1 представлена блочная схема предлагаемой формальной модели нейрона. Входы нейрона xit подаются на блоки задержки Di для задержки сигнала на время Di , а затем на триггерные элементы Ti для удлинения сигнала на величину ti . Данные элементы обеспечивают некоторую неопределенность момента поступления входного сигнала по отношению к моменту генерации выходного спайка и образуют таким образом синаптическую память, поскольку входной сигнал запоминается в этих элементах на некоторое время.
С учетом задержек Di и ti получаем, что, если на выходе обученного нейрона в момент t появился единичный сигнал, то единичные импульсы на входы нейрона поступили в интервалы времени di = [ t - ti - Di ; t - Di -1]. Неопределенность моментов поступления входных импульсов будет тем меньше, чем меньше задержки ti.
Пример временной диаграммы работы обученного нейрона с двумя входами и с заданными задержками Di и ti иллюстрирует рис. 3.2.2. Вопросительными знаками показаны неопределенности моментов прихода входных импульсов, соответствующие интервалам di.
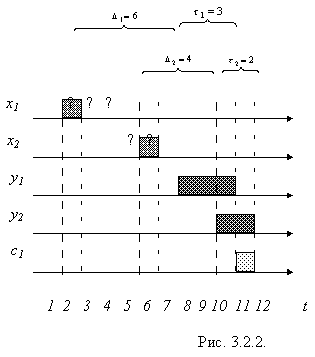
Различное отношение и расположение задержек Di и ti во времени наделяет нейрон возможностями формирования и распознавания образов следующих видов.
Если ![]() , то имеем пространственный образ. Например, образ некоторой геометрической фигуры.
, то имеем пространственный образ. Например, образ некоторой геометрической фигуры.
Если ![]() , то имеем образ следования (важен порядок следования образующих, допустима неопределенность в конкретных моментах прихода импульсов). Примером может быть распознавание слов при чтении по буквам.
, то имеем образ следования (важен порядок следования образующих, допустима неопределенность в конкретных моментах прихода импульсов). Примером может быть распознавание слов при чтении по буквам.
Если "ti = 1, то имеем пространственно-временной образ (ПВО), в этом случае однозначно определено, по каким входам и в какие моменты времени приходили сигналы. Примером может быть распознавание музыкального тона определенной высоты.
Элемент lw предназначен для набора статистики по данному пространственно-временному образу. Значение lw=1 указывает на то, что данный нейрон обучен.
Задержки Di и ti заданы изначально, т.е. являются константными параметрами нейрона. Если поступающая на данный нейрон последовательность сигналов содержит закономерность, описываемую такими временными параметрами, то нейрон сформирует образ такого пространственно-временного прообраза. Очевидно, что необходимое число нейронов такого типа в сети будет тем меньше, чем больше априорной информации о временных характеристиках прообразов известно на стадии синтеза сети.
Приведем формализм нейрона.
Owt+1 = cwt+1 & Swt+1 ;
cwt+1 = bwt+1 & lwt+1 ;
bwt+1 = 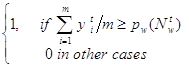
yit = ù cwt & ait ;
ait = ![]() ;
;
где ![]()
![]() = tï xit = 1 - момент импульса на входе xi ;
= tï xit = 1 - момент импульса на входе xi ;
lwt = 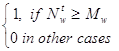 ;
;
Nw0 = 0;
Nwt = Nwt-1 + bwt ;
Rw(Nwt) - убывающая сигмоидальная функция.
Похожие работы
... , бистабильность восприятия. В дальнейшем планируется разработка программных моделей более сложных нейронных сетей и их комбинаций с целью получения наиболее эффективных алгоритмов для задачи распознавания образов. Литераура. 1.Горбань А.Н.,Россиев Д.А..Нейронные сети на персональном компьюере. 2. Минский М.Л.,Пайперт С..Персепроны.М.: Мир.1971 3. Розенблатт Ф.Принципы ...
... того привлекает огромный запас контекстных знаний, который системам распознавания образов пока недоступен. 7. Заключение Дан обзор различных нейросетевых методов распознавания изображений. Рассмотрены достоинства и недостатки этих методов при распознавании двумерных и трёхмерных объектов. Указаны проблемы при распознавании трёхмерных объектов. Выделены перспективные направления в распознавании ...
... в популяциях, которые являются существенными для развития. Точный ответ на вопрос: какие биологические процессы существенны для развития, и какие нет? - все еще открыт для исследователей. Реализация генетических алгоритмов В природе особи в популяции конкурируют друг с другом за различные ресурсы, такие, например, как пища или вода. Кроме того, члены популяции одного вида часто конкурируют ...
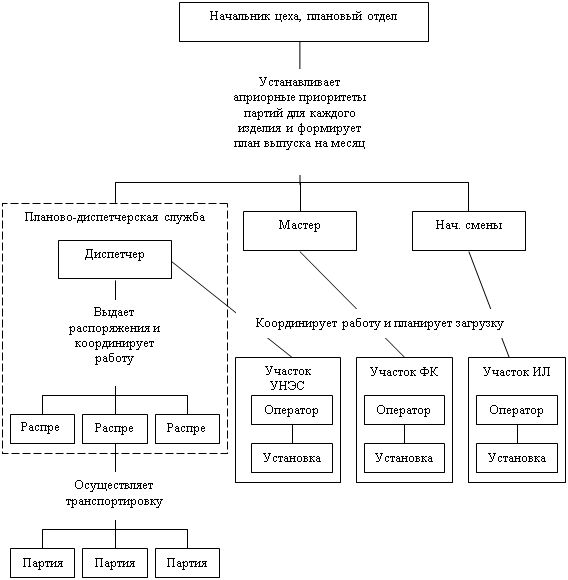
... на дипломное проектирование. Необходимо разработать программу регистрации процеса производства партий полупроводниковых пластин для использования в автоматизированной системе управления. Программа должна обеспечивать контроль и регистрацию производственного процесса производства партий пластин. Вести учет за прохождением партий полупроводниковых пластин по технологическому маршруту. Разработку ...
0 комментариев