Определение величины работы, совершаемой в системе
Если k > 0
Ионное взаимодействие
Гидрофобные взаимодействия
Вращательная диффузия К=0,34 сек
Оба отсека по отдельности должны быть электронейтральными, то есть в каждом отсеке число "+" ионов должно быть равно числу "–" ионов
Связывание проникающей молекулы с определенным участком внутри канала или вблизи него
Эти виды ионной проницаемости обнаруживают самостоятельную изменчивость в зависимости от функционального состояния клетки
Навигация
Если k > 0
Конспект лекций по биофизике
92536
знаков
0
таблиц
0
изображений
1. Если k > 0
t → ∞, N(t) → ∞ растущая
2. Если k < 0 (k2 < k1)
t → ∞, N(t) → 0 вымирающая
3. Если k = 0 (k2 = k1)
t → ∞, N = N0 cтационарная
Как изменится количество клеток в системе, если ограничить количество питательных веществ?
В этом случае изменение количества клеток в популяции со временем будет описываться логистическим уравненем Ферхюста:
dN/dt = kN*(Nmax–N/Nmax)
Nmax – максимально возможная численность популяции в данных условиях.
Рисунок. Логистическая кривая.
Начальная часть N << Nmax экспененциальный рост,
Вторая часть – изгиб в другую сторону N → Nmax количество питательных веществ ограничивает дальнейший рост количества клеток в популяции.
Основные особенности кинетики биопроцессов1. В биокинетике в качестве переменных величин выступают не только концентрации веществ, но и другие параметры.
2. Биосистема пространственно гетерогенна, следовательно условия действия реагентов могут различаться в разных точках системы и переменные изменяются не только во времени, но и в пространстве.
3. Существуют специфические механизмы саморегуляции действия по принципу обратной связи.
4. Трудности биокинетики связаны так же с тем, что она описывает процессы открытых систем.
Схема системы с отрицательной обратной связьюОУ – объект управления,
РВ – регулируемая величина,
ИУ – измерительное устройство (измерение параметров регулируемой величины)
АС – аппарат сравнения,
ОС – обратная связь,
f – сигнал от высших центров регуляции.
Простейшая кинетическая модель открытой системы. Модель системы в которой происходит обмен веществ "а" и "b" с окружающей средой, внутри обратимые реакции превращения "а" в "b", во внешних резервуарах концентрация этих веществ постоянна и равна соответственно А и В.
da/dt = k1(A–a)–k2(a–k–2b)
db/dt = k2a–k3(b–B)–K–2b
Для стационарного состояния будет соблюдаться условие: da/dt = 0, db/dt = 0.
"а" стационарное и "b" стационарное не зависят от начальных условий, то есть от значений "а" и "b" в момент t = 0. "а" стационарное и "b" стационарное определяются только величинами констант k с 1 по 3 и концентраций веществ во внешних резервуарах системы, то есть А и В.
Вывод:
В каком бы начальном состоянии ни находилась система, в ней в конце концов установится один и тот же стационарный режим при котором а = а стационарное, b = b стационарное. Это свойство эквивалентности стационарных состояний. Оно присуще открытым системам и постоянно встречается при изучении свойств биополимеров.
Качественный анализ кинетической моделиОсновная идея метода заключается в отказе от нахождения точных аналитических решений диф уравнений. Вместо этого используются качественные характеристики динамического поведения системы: устойчивость или неустойчивость стационарного состояния, переходы между стационарными состояниями, наличие колебательных движений в системе, качественная зависимость поведения системы от критических значений параметров. Наиболее важным свойством стационарного состояния является его устойчивость, она определяется спосбностью системы самопроизвольно в него возвращаться после внесения внешних возмущений, отклоняющих систему от исходно стационарной точки.
Очевидно, чтобы сделать заключение об устойчивость стационарного состояния необходимо иметь соответствующие критерии.
Бассейн с водой открытая система. С определенной Vпр в него поступает вещество а, но оно с определенной Vот из системы истекает. Vпр постоянна, Vпр = V0 = cosnt. Чтобы выяснить с какой скоростью меняется количество вещества в системе, нужно вычислить: da/dt = Vпр – Vот = V0 – ka, k – const Vот.
Рисунок. Стационарное состояние в т. а отвечает условию, что V = cosnt = 0. В стационарной точке da/dt = 0. Количество вещества в системе постоянно. Качетвенный анализ дается графическим методом. Случайные отклонения а будут компенсироваться системой. Стационарное состояние а устойчиво.
Качественный критерий устойчивости стационарного состояния ЛяпуноваЕсли система находится в состоянии равновесия, то точка, изображающая местоположение исследуемого показателя на графике будет името постоянное значение координат.
dx/dt = 0, dy/dt = 0
x(t) – const, y(t) – const
Такая точка получила название особой точки. Она показывает местоположение на графике стационарной системы. Если система по каким то причинам выходит из состояния равновесия, то изображающая точка сместится из особой точки и начнет двигаться по плоскости в соответствии с изменением координат х и у.
В этой ситуации: dx/dt = p; p = f (x;y); dy/dt = q; q = f (x;y).
p и q – непрерывные функции, определенные в данной области плоскости. В соответствии с критерием Липунова состояние равновесия устойчиво, если для любой области допустимых отклонений от состояния равновесия (e) можно указать область d, окружающую состояние равновесия и обладающую тем свойством, что ни одно движение преображающей точки, начинающееся в пределах области d никогда не достигнет границ области e.
При этих условиях стационарное состояние устойчиво.
Если же для какой то области e не существует области d, то равновесие не устойчиво.
Во многих системах существует не одно, а несколько стационарных состояний, свойства их чаще всего различаются. И это в первую очередь касается их устойчивости, поэтому в данных ситуациях задачей качественного анализа является определение устойчивости всех стационарных состояний и условий перехода между ними.
Редукция числа уравнений. Принцип узкого местаЖелательно отразить в системе уравнений все ее наиболее значимые свойства. Но вместе с тем системы диф уравнений из большого их числа, являются перегруженными. Такая модель чересчур детализирована, следовательно наиболее оптимальными моделями, характеризующими основные свойства систем являются модели, состоящие из небольшого числа диф уравнений (предположительно из двух).
Принцип узкого места (ПУМ) основан на разделении всех переменных, характеризующих свойства системы на быстрые и медленные. Характерное время процесса – t отражает время развития процесса. t процессов ферментативного катализа 10–1 – 10–6 с, процессы физиологической адаптации, для них t несколько минут и больше, процессы репродукции в этой же системе, для них t несколько минут и больше. t – величина противоположная скорости. V=1/t. В пределах одной отдельной цепочки взаимосвязанных реакций всегда имеются наиболее медленные и наиболее быстрые стадии.
Согласно ПУМ общая скорость всей цепи реакций определяется наиболее медленной стадией (она и есть узкое место), она имеет самое большое t, Vmin. Общее время всей цепи реакций (всего процесса) будет мало отличаться от характерного времени узкого места. Чтобы воздействовать на время процесса нужно воздействовать на узкое место.
При внешних возмущениях в системе наблюдаются изменения как быстрых, так и медленных перменных, однако эти изменения протекают с разной скоростью. В устойчивой системе быстрые переменные быстро отклоняются от своих начальных значений, но быстро в них возвращаются. Медленные переменные изменяются в ходе длительных переходных процессов, определяющих динамику всей системы. Фактически быстрые переменные колеблются возле своих стационарных значений. Поэтому вместо диф уравнения, описывающего динамику быстрой переменной можно записать алгебраическое уравнение, отражающее ее стационарное значение, что приведет к постоянному уменьшению числа диф уравнений в системе, останутся лишь те, что описывают наиболее медленные процессы.
dx/dt=AF(x;y)
dy/dt=Q(x;y)
A>>1 Þ A*F >> 1 Þ x быстрая переменная (dx/dt быстрая величина, скорость D х велика)
делить на А
e(dx/dt)=F(x;y), где e=1/A, e<<1, e®0, F(x;y)=0, e=0
Следовательно у является управляющим параметром, влияющим на координаты в стационарной точке. В био системах роль узкого места могут выполнять разные звенья цепи в зависимости от условий.
Например, ф/с:
РисунокВ данном процессе меняется управляющая стадия в зависимости от освещения. При плохом освещении узким место ф\с-а являются начальные фотохимические стадии поглощения и трансформации энергии и света в пигментном аппарате. Скорость этих процессов не зависит от t0 в промежутке от +5 до +300 С. При хорошем освещении узким местом ф\с-а являются темновые процессы переноса электрона и поглощения воды.
Эти процессы не справляются с потоком электронов, поступающих от пигментного комплекса, что приводит к насыщению ф\с-а (световое насыщение), эти процессы являются ферментативными, поэтому их скорость зависит от t0. И скорость ф\с-а будет увеличиваться с ростом t0.
Типы динамического поведения био систем
Система двух диф уравнений, модель хар-ся отсутствием перегруженности, на их основании можно качественно провести анализ.
dx/dt=P(x;y)
dy/dt=Q(x;y)
Используется метод фазовой плоскости
Фазовая плоскость – это плоскость с осями координат, на которых отложено значение переменных (х;у), отражающих состояние системы, таким образом каждая точка этой плоскости будет соответствовать определенному состоянию системы
х0, у0 – начальные состояния системы.
Траектория из последовательности точек, каждая из которых будет характеризовать состояние системы в любой определенный момент времени.
Последоват. сов-ть точек на фазовой плоскости, отражающая значение переменных (х;у) на пути перехода – это линия, получившая название фаз???
Изображающая точка – точка на фазовой плоскости, отражает состояние системы в определенный момент времени. Фазовый портрет – совокупность фазовых траекторий, отражающих качественные черты поведения системы во времени. P(x;y)=0 –Q(x;y)=0 –
стационарное состояние
Для нахождения особой (стационарной) точки, необходимо построить на фазовой плоскости кривые P(x;y)=0; Q(x;y)=0. Очевидно, особая точка будет находиться в месте пересечения этих кривых.
dx/dt=k1A – k1x+k2y-kx=P(x,y)
dy/dt=k2x-k-2y-k3y+k3B=Q(x,y)
y=-C1x+C2
y=C3x+C4
C – коэффициент пропорциональности
Графики могут пересекаться в нескольких точках (если это кривые), следовательно существует несколько стационарных состояний.
Фазовый портрет триггерной системы
Типы устойчивости особых точекВажной задачей является определение устойчивости особых точек. Производится по виду правых частей исходной системы уравнений. Об устойчивости стационарного состояния системы судят по поведению системы в случае небольшого отклонения от стационарной точки.
e=x-xст
h=у-уст
Для определения характера устойчивости необходимо одновременно учитывать поведение во времени отклонений e и h. Существуют специальные уравнений, описывающие e и h.
e(t)=C11el1t+C12el2t
h(t)=C21el1t+ C22el2t
Особый смысл имеют l1 и l2 – это экспоненциальные показатели
l1,2 = ![]()
a,b,c,d – значения частных производных в точке (хстац;устац). От вида l1,2 зависит поведение отклонений e и h соответствующих поведению х и у в особой точке (окресностях). l1,2 это либо действительные числа, либо комплексно-сопряженные (если под знаком корня дробь).
1. l1 и l2 < 0 то есть они являются действительными отрицательными числами, значение e и h будут со временем снижаться, то есть отклонение системы от особых точек со временем будет . В этом случае стационарное состояние является устойчивым, а особая точка называется устойчивый узел, такой точке соотвествует особый тип фазового портрета.
Рисунок. Система будет возвращаться по какой-то траектории в стационарное состояние.
2. l1 и l2 > 0, действительные положительные числа e и h будут увеличиваться со временем, следовательно первоначальное состояние было неустойчиво и система все дальше будет отклоняться от состояния равновесия.
Неустойчивый узел. Фазовый портрет такой же, но стрелки на периферию.
3. l1 и l2 действительные числа разных знаков.
Рисунок. Тогда на фазовом портрете системы будет существовать особая точка типа "седла". Сопаратиссы.
Из любого начального положения на фазовой плоскости кроме особой точки сепаратисс система будет удаляться из стационарного состояния. Если l1 и l2 комплексно-сопряженные числа, то изменения во времени e и h носят колебательный характер. Частные случаи:
1. Действительные l1 и l2 < 0,
Рисунок. Re<0, то колебания ситемы носят затухающий характер. Особая точка на фазовом портрете будет называться устойчивый фокус.
2. Действит l1 и l2 > 0,
Рисунок. Cтрелки на фазовом портрете направлены наружу, неустойчивый фокус
3. Re l1 и l2 = 0,
Рисунок. В этом случае l1 и l2 превращаеются в мнимые числа, фазовые траектории будут представлять собой эллипсы, не проходящие через начало координат. В начале координат находится неустойчивая точка (центр). Необольшие возмущения в системе переводят ее с одной траектории на другую, то есть изменяется амплитуда колебания.
Первые пять типов состояния равновесия являются грубыми, так как их характер не изменяется существенно при небольших изменениях правых частей исходного уравнения, а так же из проиводных первого порядка. Эти типы устойчивости характерны для био систем, так как они должны определенным запасом грубости. Такой запас позволяет им сохранить основные динамические свойства при умеренных внешних воздействиях.
Биологические триггеры(Триггер – спусковой крючок у оружия)
Любая триггерная система способна переключаться с одного режима на другой. Ф.п. стриггер системы характризуются как минимум двумя стационарными точками (А и С)
Рисунок. 2 вида переключения.
1) силовое переключение осуществляется при значительном изменении переменных х или у.
3) связан с наличием управляющего параметра. Он оказывает влияние на величину обеих переменных х и у.
После этого можно восстановить значение управляющего парметра, что приведет к восстановлению исходного вида фазового портрета, однако система при этом остается в устойчивой точке С.
Колебательные процессыхарактерная черта био систем. Частые колебания промежуточных продуктов био-хим реакций, численности видов, потенциала мембраны и т.д.
В любом организме существует набор био-хим процессов, в их основе лежат внутренние свойства системы. Причины колебаний во внутренних динамических свойствах системы. Автоколебательные процессы. Тип фазового портрета – устойчивый предельный цикл. Замкнутая траектория, не проходящая через начало координат. С наружной и внутренней стороны подходят спиральные траектории. Система работает в устойчивом колебательном режиме. Если система в силу внешних воздействий выйдет из такого режима, то в силу своих свойств она вернется на замкнутую траекторию. Возвращение будет осуществляться по спиральной траектории. Переход движения вдоль траектории предельного цикла в автоколебательной системе не зависит от начальных условий.
Распределение биологической системы
В био кинетике предполагается изменение переменных не только во времени но и в пространстве, в биохимии важнее изменения во времени. В отличие от точечные, такие модели, которые учитывают измениения переменных во времени и в пространстве, называются распределенными. Таким образом, в распределенных системах будут 2 параллельных процесса – хим превращения в отдельных точках пространства и процесс диффузии веществ из области высокой в область низкой концентрации. То есть происходит перенос вещества между соседними элементарными отсеками. В реальных био системах часто существует пространственное распределение источников энергии. Эти системы называются активные распределенные системы.
Анализ распределенной системы:
Состояние системы уравнений, отражающих хим реакцию и диффузию реагента. Max простой пример распределенной системы – система, в которой имеется одна переменная Х, одновременно участвующая в хим процессе и диффунцирующая вдоль узкой трубки. r – учитывается размер трубки. Диффузионный поток вещества – масса вещества, проходящего через единицу площади перпендикулярной к направлению диффузии (ось х), пропорционален градиенту вещества, взятому с обратным знаком.
I = –D*ds(r,t)/dr,
D – коэффициент диффузии, t – время, C – концентрация, изменение концентрации во времени за счет диффузии в элементарном объеме трубки, заключенном между точками r и r+Dr, зависит от разности потоков в точках r и r+Dr. Если D const, то изменение С во времени (скорость изменения С) =
dc/dt=D*d2c(r,t)/dr2,
уравнение диффузии, оно описывает скорость изменения С в системе, в которой происходит только диффузия вещества. Но по условию, в распределенной системе параллельно происходит и хим превращение вещества. Величиной, отражающей хим превращение в системе, является величина точечных членов, которая обозначается, как функция концентрации.
dc/dt=f(c)+D*d2c(r,t)/dr2,
выражение отражает химическое превращение и изменение концентрации. В этой системе изменения происходят только по оси Х. В системах, в которых происходят колебательные процессы невозможно отразить их поведение с помощью одного уравнения, поэтому в данном случае используются базовые модели с двумя переменными:
dx/dt=P(x,y)+Dx*d2x/dr2,
dy/dt=Q(x,y)+Dy*d2y/dr2
Такая базовая модель позволяет качественно описать процессы самопроизвольного возникновения волн и структур в распределенных системах. В общем, они называются самоорганизацией, она возникает, когда в системе появляется неустойчивость, приводящая к потере исходного распределения веществ во времени и пространстве. Вместо этого устанавливается новое распределение вещство во времени и пространстве. Характер самоорганизации зависит от функции f (P,Q) (точечных членов) и D, в частности существуют следующие виды самоорганизации:
1. Распределение возмущения в виде бегущего импульса (нервный импульс).
2. Стоячие волны.
3. Синхронные автоколебания разных элементов во всем пространстве системы.
4. Стационарные неоднородные распределения переменных в пространстве (диссипативные структуры).
5. Генерация волн автономными источниками импульсной активности (локальные кратковременные флуктуации переменных).
Общим условием возникновения процессов самоорганизации всегда является появление неустойчивости в исходной распределенной системе.
Молекулярная биофизика
Предмет молекулярной биофизикиИзучает строение и физ хим свойства биофункциональных молекул (прежде всего биополимеров). Основной задачей мол биофизики является вскрытие физических механизмов, ответственных за био функциональность этих молекул (напимер, ферментативная активность белков).
Методы: ЭПР, ЯМР, рентгеноструктурный анализ, биохимические технологии.
Стремится выяснить основные детали структуры и функции молекул. Основное свойсто – хиральность биомолекул. Большинство молекул, содержащих больше 3х атомов, не имеют ни центра, ни плоскости симметрии. Их можно назвать диссиметричные, или хиральные. В био молекулах связи С могут быть заняты, как одинаковыми, так и разными группами.
СХ2НZ молекула будет иметь плоскость симметрии. Рисунок
Из 20 а-к хиральность свойственна 19, ей не обладает глицин. В процессе хим синтеза из исходных симметричных молекул хиральное вещество образуется всегда в виде рацимической смеси, которое содержит по 50% молекул D, L – форм хирального вещества. Состояние с максимальной энтропией.
В живой природе имеется фиксация в био структурах какой-либо одной формы хиральных веществ (например в ДНК и РНК всегда D-форма углеводов). С позиции биофизики это объясняется фиксацией информации, то есть выбор одного антипода равнозначен информации в 1 бит.
L-аспарагиновая кислота не имеет вкуса, D-аспарагиновая кислота сладкая. Существуют некоторые яды, вызывающие токсический эффект в одной форме и безвредные в другой. Это свойство открыто в 1848 г. Л. Пастером.
Виды взаимодействий биологических молекулОсновой структуры биологических молекул являются сильные связи – химические ковалентные связи. Биомолекулы отличаются высоким содержанием С, между С–С сильная связь, энергия разрыва равна 328,9 кДж/моль. Сильные связи присутствуют там, где содержатся мономерные звенья. Сильные связи образуются внешними электронами атомов, поэтому для исследования их особенностей используется раздел Квантовая Химия. Но есть важный недостаток сильной связи, они создают жесткий каркас.
Слабые связи основаны на действии невалентных слабых сил, и на их базе формируются биомолекулы. Различаются взаимодействия белковых молекул с другими видами молекул. На базе слабых связей осуществляется тонкая регуляция химических взаимодействий, компартментализация, градиент.
Слабые связи характеризуются преобладанием сил притяжения на больших расстояниях и преобладанием сил отталкивания на близких расстояниях.
Рисунок 1
График зависимости потенциальной энергии слабого взаимодействия от расстояния между двумя взаимодействующими частицами молекулярной природы.
R0 – минимальный потенциал энергии взаимодействия. В точке, соответствующей R0 силы притяжения равны силам отталкивания. Если расстояние меньше R0 , то преобладают силы отталкивания. Если расстояние больше R0, то преобладают силы притяжения. R0 оптимальное расстояние, на котором и будут находиться взаимодействующие частицы.
Вторичная структура биомолекул зависит от различных видов слабых связей.
Похожие работы
... Во времена, например, Платона. Они были одни, а на современном этапе они другие. По мнению Карпекова К.С.(2).(который по моему мнению наиболее полно обозначил современную концепцию происхождения объектов Вселенной.). В.И. Вернадский впервые поставил вопрос о рациональном контроле над окружающей средой. Ноосфера Вернадского – это сфера материального, в котором в качестве управляющего, регулирующего ...
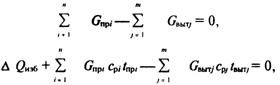
... или технологических процессов; – при выборе технического решения обеспечить малоотходность производства и максимальную эффективность использования энергоресурсов. Задачи специалиста в области безопасности жизнедеятельности сводятся к следующему; – контроль и поддержание допустимых условий (параметры микроклимата, освещение и др.) жизнедеятельности человека в техносфере; – идентификация ...
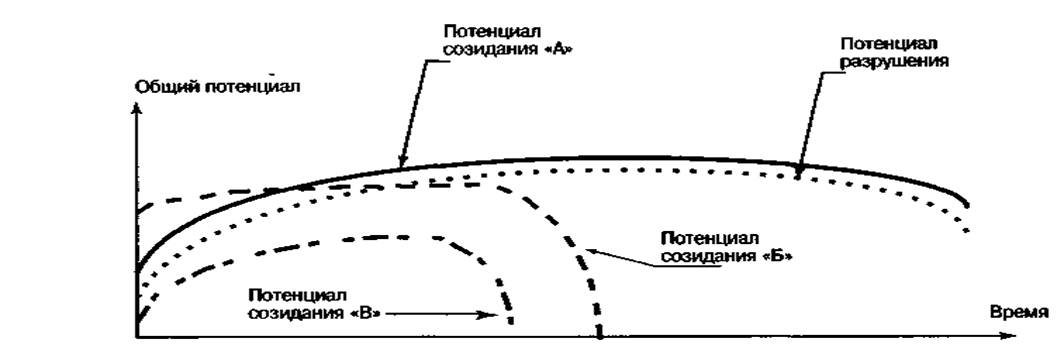

... не знают о законе. Второй вариант. Руководитель знает о законе, а его подчиненные нет. Третий вариант. Руководитель и подчиненные знают о законе», единства анализа и синтеза Список использованной литературы 1. Алиев В.Г., Варфоломеев В.П. Теория организации, курс лекций. «Самоорганизация в природе и обществе», Махачкала ИПЦ ДГУ, 1998. 2. Доблаев В.Л. Теория организаций. — М.: Институт ...
... не тепловая энергия. Вероятно, для выяснения истины необходимо более широкое, чем термодинамическое или статистическое, определение энтропии. Поэтому-то мы рассматриваем этот вопрос здесь, в разделе биологических концепций. Энтропия и эволюция. Вероятно, зарождение проблемы взаимосвязи между энтропией и эволюцией произошло в 1854 г., когда Гельмгольц и Больцман первыми обратили внимание на ...
0 комментариев