Определение величины работы, совершаемой в системе
Если k > 0
Ионное взаимодействие
Гидрофобные взаимодействия
Вращательная диффузия К=0,34 сек
Оба отсека по отдельности должны быть электронейтральными, то есть в каждом отсеке число "+" ионов должно быть равно числу "–" ионов
Связывание проникающей молекулы с определенным участком внутри канала или вблизи него
Эти виды ионной проницаемости обнаруживают самостоятельную изменчивость в зависимости от функционального состояния клетки
Навигация
Связывание проникающей молекулы с определенным участком внутри канала или вблизи него
Конспект лекций по биофизике
92536
знаков
0
таблиц
0
изображений
1. Связывание проникающей молекулы с определенным участком внутри канала или вблизи него.
2. Основная причина – транспорт вещества через мембрану с помощью молекулы-переносчика:
а) количество молекул-переносчиков ограничено,
б) скорость из реагирования с переносимым веществом так же лимитирована.
Скорость облегченной диффузии достигает max, когда все молекулы переносчика будут заняты транспортируемым веществом.
Теория облегченной диффузии напоминает теорию субстрат-связывающего комплекса. Данный вид транспорта можно ингибировать с помощью химических аналогов транспортируемого вещества.
Механизмы первично активного транспорта
Энергия клеточного механизма концентрируется в виде АТФ. Существуют специальные мембранные насосы, их совокупность – первично активный транспорт. Источник энергии – клеточный метаболизм, если отключить источник энергии, то ионы расположатся равномерно, относительно мембраны. Концентрационный градиент направлен внутрь клетки, ионы Na пассивно поступают внутрь клетки. Но концентрационный градиент постоянен, так как ему противостоят Na насосы.
Основные особенности первично активного транспорта:
1. Осуществляется против концентрационного градиента.
2. Система первичного транспорта в высшей степени специфична (Na система не перекачивает другие ионы).
3. Для его обеспечения необходима АТФ или другие источники энергии (метаболические яды блокируют насос).
4. Обменивает один вид ионов на другой (К-Na насос).
5. Многие виды ионных насосов выполняют электрическую работу, перенося заряды через мембрану (реогенный насос – это насос, при работе которого создается электрический ток).
6. Активный транспорт с помощью ионных насосов избирательно подавляется блокирующими агентами. (Существуют специфические вещества, которые блокируют данный насос, например, убаин – сердечный гликозид. Это вещество конкурентно блокирует участки, связывающие ионы К+.)
7. Энергия, необходимая для первично активного транспорта, высвобождается при гидролизе АТФ ферментами, расположенными в мембране. Активность ферментов зависит от концентрации ионов.
Современная гипотеза первично активного транспорта
K-Na-АТФаза – молекула из двух субъединиц, имеющих внутренние полости: a-большая субъединица (полипептид), b-малая субъединица (гликопротеид). a обладает высоким сродством к Na +, b - к К+. Полость a-субъединицы заполняется 3 ионами Na+, полость b-субъединицы заполняется 2 ионами К+. Потом у a-субъединицы сродство к Na+ падает, а у b-субъединицы сродство к К+ возрастает.
За счет флуктуации происходит пространственное совмещение полостей субъединиц и обмен ионами. В конце цикла полости открываются, и ионы их покидают.
Другая гипотеза. В начале происходит заполнение полостей описанным выше способом, затем поворот K-Na-АТФазы на 1800. После чего ионы покидают полости, а K-Na-АТФаза опять поворачивается на 1800. Если молеула постоянно переворачивается, то это должно привести к перестройке молекулярного слоя – спорный момент.
Механизм вторично активного транспорта
заключается в переносе веществ через мембрану против концентрационного градиента, обеспечиваемом энергией, которая высвобождается при переносе другого вещества по градиенту. То, что транспортируется по градиенту, называется синпортом, или ко-транспорт. Пример, транспорт а-к или сахаров через био мембраны.
Транспорт в клетки аланина.
В присутствии внеклеточных ионов Na+ транспорт аланина в клетки осуществляется до тех пор, пока внутриклеточная концентрация Na+ будет в 7-10 раз больше внеклеточной. Если во внеклеточной среде Na+ отсутствует, то концентрация аланина внутри клетки не отличается от внеклеточной.
2 рисунка. max скорость транспорта в двух случаях одинакова. Внеклеточный Na+ оказывает непосредственное влияние на транспорт аланина (различный наклон графиков). Если повысить внутриклеточную концентрацию Na+, то аланин из клетки будет выходить во внеклеточную среду. Вторично активный транспорт не зависит от концентрации Na+ вне клетки, а зависит от концентрации градиента ионов Na+. Градиент Na+ является движущей силой, промежуточной стадией в процессе использования энергии (в системе вторично активного транспорта).
Системы антипорта, или контр-транспорта – это система вторично активного транспорта, функциорующая на основе переносчика обменника, обеспечивающего выведение из клетки транспортирующего вещества против его концентрационного градиента в обмен на сопряженный, пассивно поступающий в клетки поток ионов Na+. Движущей силой является потенциальная энергия концентрационного градиента ионов Na+.
Примеры:
1. Транспорт Ca2+ в обмена на Na+, осуществляется во многих типах клеток. Уровень концентрации внутриклеточного Ca2+ 10–9 моля, внеклеточной – 10–6 моля. Мышечное сокращение, выделение нейромедиаторов синапсами регулируется Са2+-зависимыми К+-каналами. Механизм: и ионы Nа+ и ионы Са2+ имеют один и тот же переносчик, у внутренней поверхности мембраны, обладающей высоким сродством к Са2+, у наружной – к Nа+. Са2+-насос удаляет Са2+ из клетки.
2. Антипортная система клеток проксимальных отделов нефрона, обменивающая Nа+ на Н+. Из мочи к клетки проксимальных отделов нефрона выделяется Nа+, взамен выводится Н+. Система не совершает электрическую работу, поэтому не надо тратить энергию. K-Na-насос сохраняет Nа+ в организме.
Визикулярный транспорт
происходит путем эндо- и экзоцитоза. Это вид транспорта, при котором вещества перекачиваются внутрь клетки или из нее внутри маленьких пузырьков или визикул. Жидкие вещества – пиноцитоз, твердые вещества – фагоцитоз. Когда переносятся гормоны или медиаторы, то вначале они взаимодействуют с мембранными рецепторами. Рецепторы обладают способностью к латеральной диффузии, при этом образование комплекса рецептора с лигандом вызывает перемещение этого комплекса в углубление мембраны.
Образуется окаймленная ямка. Внутренняя поверхность покрыта особым белком – клатрином, он связывает занятую лигандом молекулу рецептора, затем этот белок участвует в отшнуровывании визикулы от поверхности мембраны. Визикулы разрушаются, но некоторые проходят насквозь, и тогда вещества высвобождаются с другой стороны. Когда визикула разрушается внутри клетки – эндоцитоз, вне клетки – экзоцитоз. Мембранный материал вновь включается в клетку Þ круговорот.
Потенциал покоя
это разница потенциалов между цитоплазмой и внеклеточной средой, существующая в каждой живой клетке, находящейся в состоянии покоя. Внутриклеточная среда заряжена отрицательно. Величина ПП может быть различной, зависит от состояния клетки от –15 до –90 мВ у большинства клеток.
Для того, чтобы измерить величину ПП используется микроэлектродная техника. Используется специальный вид электродов, отличающийся намного меньшим диаметром кончика (доли мкм, или 1 мкм). Бывают стеклянные и металлические, по форме напоминают копье, необходим раствор электролита для хорошего проведения электрического тока.
Основные теории ПП:
1. В 1848 г. Дюбуа-Реймон "теория электромоторных молекул" (теория заряженных диполей). Дюбуа-Реймон предположил, что на мембране находятся диполи, ориентированные отрицательным зарядом внутрь. При возбуждении диполи поворачиваются на 1800, что приводит к положительному заряду внутри клетки. Разность потенциалов в этой теории является предшествующей.
2. В 60-е годы XIX в. – альтернативная теория Германа. В состоянии покоя мембрана клетки не заряжена, и ПП отсутствует. Однако при повреждении, в поврежденном участке появляется избыток отрицательного заряда, в силу наличия кислых продуктов. Поэтому между поврежденными и неповреждеными участками возникает электрический ток. Теория Германа исключала существование разности потенциалов на мембране.
3. Теория полупроницаемой мембраны Берштейна. 1906 г. Положения:
~ мембрана обладает свойством полупроницаемости (в специальных экспериментах в группе Пфейффера показали: при пропускании электрического тока – поляризация мембраны и появление концентрационной электро-двигательной силы.
~ наличие концентрационных градиентов на биологической мембране. В исходном состоянии К+, Сl–, Na+ и другие распределены различно на мембране. Берштейн обратил внимание на К+-концентрационный градиент по направлению из клетки наружу.
Существуют механические насосы. Перемещение из клетки через мембрану осуществляется по градиенту; но выход не бесконечен, как только К+ выйдет из клетки, образуются силы, противодействующие этому выходу. 1. Первый положительный заряд на мембране препятствует выходу остальных (электростатическое отталкивание). 2. Крупные молекулы, которые не могут проходить через мембрану, мембраны их не пропускают, они не будут пропускать К+ мембранный слой ионов К+ снаружи, слой анионов изнутри. При повреждении мембраны анионы выходят наружу и создается электрический ток.
Еm = RT/F * ln([K+]ant/[K+]in)
Еm – ПП, F- число Фарадея.
Эксперимент: кнаружи поверхности мембраны облицеров. соли К+ Þ выход ионов К+ из клетки затруднен Þ величина ПП?
Результат: ампликация солей К+ к наружной поверхности мембраны приводит к снижению величины ПП, степень снижения оказалась пропорциональной концентрации К+ в облицеров. растворе.
Вывод: К+ имеет основное значение в генерации ПП.
Если удалить Na+ или изменить его концентрацию во внеклеточной жидкости, это не толко не снизит величину ПП, но даже приведет к небольшой дополнительной поляризации мембраны Na+ не играет существенной роли в генерации ПП.
Прямые определения концентрации К+ в клетке и окружающей среде показали хорошее соотношение с теоретически рассчитанными значениями ПП. После Бернштейна данные подтверждались.
Джерард и Фурузава, 1930 г. Работали на нервах краба в условиях гипоксии и апоксии (полное отсутствие О2). ПП прогрессивно снижался и, наконец, исчез в нормальном целостном участке нерва.
Вывод: энергия, необходимая для генерации ПП берется из окислительных процессов.
Современная мембранная теория
Ходжкин и Хаксли разработали в 30х годах ХХ в., используя микроэлектродную технику на гигантских аксонах кальмара. ПП –50 мВ. При возбуждении генерации ПД амплитуда до 100 мВ.
С позиции теории Берштейна. При возбужедении растет проницаемость мембраны для всех ионов. При этом происходит перераспределение этих ионов в сторону выравнивания концентрации внутри и вне клетки, поэтому ПД = ПП (» –50 мВ) Þ кризис мембранной теории Берштейна.
Основные положения современной мембранной теории
1. Клеточные мембраны обладают избирательной проницаемостью не только к ионам состоянии К+, но и к Na+ и к другим.
Похожие работы
... Во времена, например, Платона. Они были одни, а на современном этапе они другие. По мнению Карпекова К.С.(2).(который по моему мнению наиболее полно обозначил современную концепцию происхождения объектов Вселенной.). В.И. Вернадский впервые поставил вопрос о рациональном контроле над окружающей средой. Ноосфера Вернадского – это сфера материального, в котором в качестве управляющего, регулирующего ...
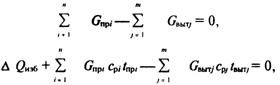
... или технологических процессов; – при выборе технического решения обеспечить малоотходность производства и максимальную эффективность использования энергоресурсов. Задачи специалиста в области безопасности жизнедеятельности сводятся к следующему; – контроль и поддержание допустимых условий (параметры микроклимата, освещение и др.) жизнедеятельности человека в техносфере; – идентификация ...
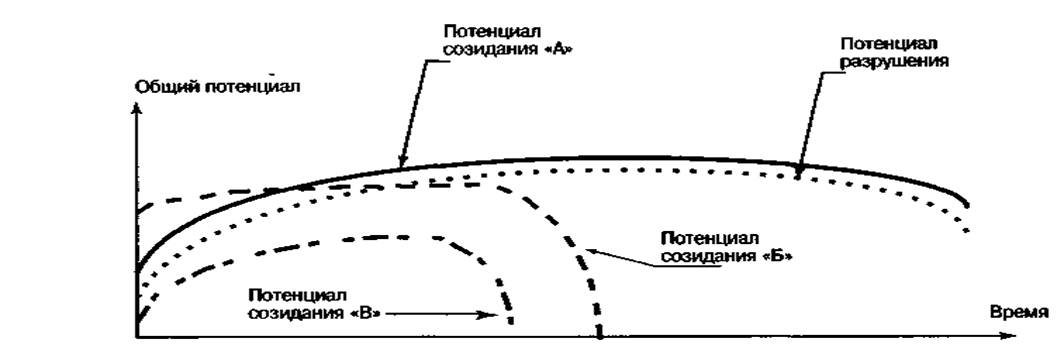

... не знают о законе. Второй вариант. Руководитель знает о законе, а его подчиненные нет. Третий вариант. Руководитель и подчиненные знают о законе», единства анализа и синтеза Список использованной литературы 1. Алиев В.Г., Варфоломеев В.П. Теория организации, курс лекций. «Самоорганизация в природе и обществе», Махачкала ИПЦ ДГУ, 1998. 2. Доблаев В.Л. Теория организаций. — М.: Институт ...
... не тепловая энергия. Вероятно, для выяснения истины необходимо более широкое, чем термодинамическое или статистическое, определение энтропии. Поэтому-то мы рассматриваем этот вопрос здесь, в разделе биологических концепций. Энтропия и эволюция. Вероятно, зарождение проблемы взаимосвязи между энтропией и эволюцией произошло в 1854 г., когда Гельмгольц и Больцман первыми обратили внимание на ...
0 комментариев