Определение величины работы, совершаемой в системе
Если k > 0
Ионное взаимодействие
Гидрофобные взаимодействия
Вращательная диффузия К=0,34 сек
Оба отсека по отдельности должны быть электронейтральными, то есть в каждом отсеке число "+" ионов должно быть равно числу "–" ионов
Связывание проникающей молекулы с определенным участком внутри канала или вблизи него
Эти виды ионной проницаемости обнаруживают самостоятельную изменчивость в зависимости от функционального состояния клетки
Навигация
Эти виды ионной проницаемости обнаруживают самостоятельную изменчивость в зависимости от функционального состояния клетки
Конспект лекций по биофизике
92536
знаков
0
таблиц
0
изображений
2. Эти виды ионной проницаемости обнаруживают самостоятельную изменчивость в зависимости от функционального состояния клетки.
В мембране диэлектриком явлется липидная фаза (конденсатор), чтобы зарядить конденсатор до –75 мВ на 1 мм2 должно находиться 5000 пар ионов. В генерации ПП участвует К+, концентрация внутри клетки в 20 раз выше Þ концентрационный градиент для внутрь клетки (так как во внеклеточной среде концентрация ионов Na+ в 5-15 раз больше, концентрация Сl– вне клетки в 20-100 раз больше.
Чем больше проницаемость мембраны для иона, тем больше в клетку вносится данного иона.
Р К+ : Р Na+ : Р Сl– = 1 : 0,04 : 0,045
Уравнение Гольдмана:
Em = RT/F * ln (PК+*[К+]out+ PNa+*[Na+]out+ PCl–*[Cl–]out)/(PК+*[К+]in+ PNa+*[Na+]in+ PCl–*[Cl–]in)
Потенцил Действия
1. Объясняется поворотом диполя на 1800.
2. Теория альтерации Германа. При возбуждении возникает избыток кислых продуктов, которые несут отрицательны заряд, что приводит к разности потенциалов между возбужденным и невозбужденым учаском.
3. Мембранная теория Берштейна. В возбужденном участке мембраны резко увеличивается проницаемость для всех ионов, концентрации ионов смешиваются и участок становится электронейтральным.
4. Ходжкин и Хаксли. Рост проницаемости мембраны для ионов в месте воздействия. При возбуждении электропроницаемость мембраны увеличивается примерно в 500 раз. Max увеличивается проницаемость мембраны для Na+ (отсюда Na-теория ПД). Na+ свободно проходит внуть клетки. При возбужедении электро-химическое равновесие определяется потенциалом Na+. Равновесный потенциал для К+ = –97 мВ, для Na+ = +50 мВ. При возбужедении мембрана перезаряжается. Положение обратной активации и инактивации Na+-каналов, Na+-канал может активироваться (открываться) при определенных значениях потенциала. Причина активации Na+-каналов – деполяризация мембраны, чем больше деполяризация, тем больше проницаемость мембраны для Na+. Зависимость близка к линейной в подкор уровне; как только мембрана достигнет критического уровня деполяризации – зависимость нелинейная, лавинообразный вход Na+ в клетку.
1). Для объяснения реполяризации используется положение об инактивации Na+-каналов. При приближении потенциала мембраны к равновесному для Na+, Na+-каналы инактивируются и посупление Na в клетку прекращается. К графику: в основе регенеративный процесс (сам себя поддерживающий), развивающийся по принципу обратной связи.
2). Рост К+ проницаемости мембраны. Не столь значителен, как для Na+ ( в 5-15 и 500 раз соответственно). Проницаемость для К+ развивается медленнее, чем для Na+. Ионы К+ в этой ситуации будут выходить наружу и выносить заряд.
3). Механизм активного транспорта, представленный K-Na-насосом. 3 Na+ внутрь и 2 К+ наружу.
Эксперименты Ходжкина и Хаксли.
Гигантский аксон кальмара. Из внеклеточной среды были удалены 2/3 Na+. При этом амплитуда ПД снизилась » на 50%. Замена внутриклеточного Na+ на другие ионы приводит к некоторому росту ПД. Замена ½ внутриклеточного К+ на Na+ приводит к значительному снижению ПД.
Метод фиксации потенциала
метод Петч-Клемпинга. С его помощью можно зафиксировать на длительное время значение мембранного потенциала на любом желаемом уровне. Это делается с помощью внешнего генератора напряжения
Суммарные мембранные токи при ПД
1. Подпороговая область:
Слабое изменение мембранного потенциала, суммарный ионный ток направлен от клетки наружу, так как поток К+, выходящий из клетки, уже усиливается из-за удаления мембранного потенциала от равновесного потенциала для К+. Входящий ток Na+ еще слаб, так как рост Na+-проницаемости пока невелик. Однако с развитием деполяризации Na-ый поток постепенно нарастает.
2. Критический уровень деполяризации:
В этот момент суммарный ионный ток через мембрану равен нулю, так как встречные токи ионов Na+ и К+ уравновешивают друг друга. Даже небольшая дальнейшая деполяризация приводит к росту входа Na+-тока в сотни раз.
3. Во время фазы деполяризации резко увеличивается Na+-проницаемость и суммарный мембранный ток, направленный внутрь клетки. Выходящий К+-ток растет медленнее и становится заметным только к моменту пика потенциала.
4. Фаза реполяризации:
В момент пика потенциала большинство Na+-каналов инактивированны, а К+-ток max. Поэтому суммарный мембранный ток – выходящий.
Кальциевая теория активации и инактивации Na+-каналов
В состоянии покоя у наружного отверстия Na+-канала находится Са2+, который электростатически тормозит проникновение Na+ в канал. При возбуждении наружная поверхность мембраны заряжена отрицательно, при этом Са2+ уходят со своих мест, вход открывается и Na+ входит в клетки.
Инактивация: по ходу деполяризации узкие Na+-каналы могут закупориваться Na+. Во многих каналах есть воротные белки (могут менять свое местоположение под влиянием изменения потенциала). В состоянии покоя активационный белок закрыт, а инактивационный открыт. При возбуждении открывается активационный белок в момент закрывания инактивационного белка. В конце реполяризации белки так же закрываются и потом открываются (исходное состояние).
Передача возбуждения по нервным волокнам
В начале 30х годов ХХ в. Хилл. 1932 г. "Химическая волна проведения в нервах". Хилл использовал разные нервы, но преимущественно краба. Даже в состоянии покоя в единицу времени вырабатывается некоторое количество тепла. Это тепло было названо теплопродукция покоя. Когда в нервном волокне возбуждение – теплопродукция возбуждения (ТВ), она делится на 2 фазы:
1. Начальная ТВ, которая составляет 2-3% от всей ТВ и приходится непосредственно на период возбуждения.
2. Задержанная ТВ » 97% всей ТВ. Если подать серийный импульс на нерв краба, то задержанную ТВ можно зафиксировать в течение 25-30 минут. Возбуждения в тканях уже нет, но ТВ имеет место.
3. Утечка тепла при работе Na.
Хилл разрабтал чувствительную теплоэлектрическую методику, которая позволяла фиксировать теплообразование в течение 20 мс. Эксперименты при О0 С. Начальную фазу теплопродукции делили на 2 периода: позитивная и негативная начальная теплопродукция. При О0 С для нерва краба позитив в начальные 20 мс = 14 мк кал. В течение последующих 150 мс » 85% тепла поглощается нервной тканью обратно (12 мк кал).
Позитивная начальная теплопродукция: причина: химические процессы, обуславливающие изменение проницаемости мембраны. При возбуждении в клетку поступает Na+ и смешивается с К+ и наоборот. Должно образовываться тепло. Это тепло покрывает до 50% позитивной начальной теплопродукции.
Негативная начальная теплопродукция: химические реакции в этот период могут быть эндотермическими. Негативная теплопродукция не является обязательной.
Проведение возбуждения
В 1885 г. Герман предложил теорию малых токов. Осуществляется последовательно между участками волокна. В участке, соседнем с возбужденным будет наблюдаться выход электрического тока.
Кабельная теория нервного волокна: нервное волокно внутри содержит проводящую среду, оболочка невного волокна имеет слой, который плохо проводит возбуждение. Нервное волокно омывается внеклеточной жидкостью, которая проводит электрический ток.
Эквивалентная электрическая схема нервного волокна
В состоянии покоя внутриклеточная среда имеет избыточный отрицательный заряд. Сила тока меняется с расстоянием от возбужденного сегмента, декремента.
Факторы, определяющие скорость распространения возбуждения по нервному волокну
1. Пространственная константа определяет величину декремента деполяризации, l - пространственная константа.
2. Коэффициент надежности, соотношение между амплитудой ПД, критической энергией и ПП. S=ПД/(Екр–ПП), ПД=120мВ, ПП=–70мВ, Екр=–55мВ Þ S=8. Чем больше S, тем быстрее проведение.
3. Временная константа t мембр. При возбуждении мембраны меняется заряд. Длительность перезарядки мембраны. tмембр=Rm*Cm. Чем больше t мембр, тем ниже С. Vраспр=S*l/tмембр.
Механизм распространения возбуждения
Возбуждение охватывает последовательно все отделы нервного волокна. R наруж влияет на скорость распространения возбуждения. В экспериментрах Ходжкина изменили внеклеточную среду на масло, которое имеет большее сопротивление, объем снизился на 30-50%. Эксперимент: нерв помещается на параллельные пластинки из серебра, замыкают с помощью ртутной ванночки, объем проведеним растет на 16-30%. Была подтверждена теория местных токов для безмякотных волокон. В мякотных нерных волокнах механизм проведения другой. Миелин имеет рост сопротивления и снижение емкости, миелиновая оболочка прерывается перехватами Ранвье – сальтаторное проведение. R на 1 см2 поверхности в перехвате Ранвье = 10-20 Ом, в миелиновой оболочке = 0,003-0,005 Ом. Петли тока в миелиновых нервных волокнах выходят через невозбужденый перехват Ранвье, находящийся спереди от возбужденного . Эксперименты Тасаки.
1. Электроды стоят на миелине, два выходящих тока (это токи, выходящие из последующего и предыдущего перехвата Ранвье. Входящий ток не регистрируется.
2. Средний электрод на перехвате. Появляется входящий ток.
Похожие работы
... Во времена, например, Платона. Они были одни, а на современном этапе они другие. По мнению Карпекова К.С.(2).(который по моему мнению наиболее полно обозначил современную концепцию происхождения объектов Вселенной.). В.И. Вернадский впервые поставил вопрос о рациональном контроле над окружающей средой. Ноосфера Вернадского – это сфера материального, в котором в качестве управляющего, регулирующего ...
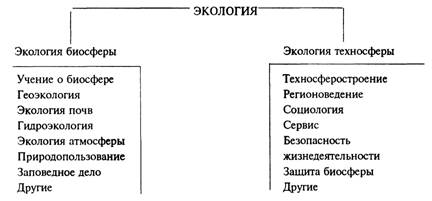
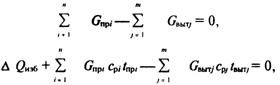
... или технологических процессов; – при выборе технического решения обеспечить малоотходность производства и максимальную эффективность использования энергоресурсов. Задачи специалиста в области безопасности жизнедеятельности сводятся к следующему; – контроль и поддержание допустимых условий (параметры микроклимата, освещение и др.) жизнедеятельности человека в техносфере; – идентификация ...
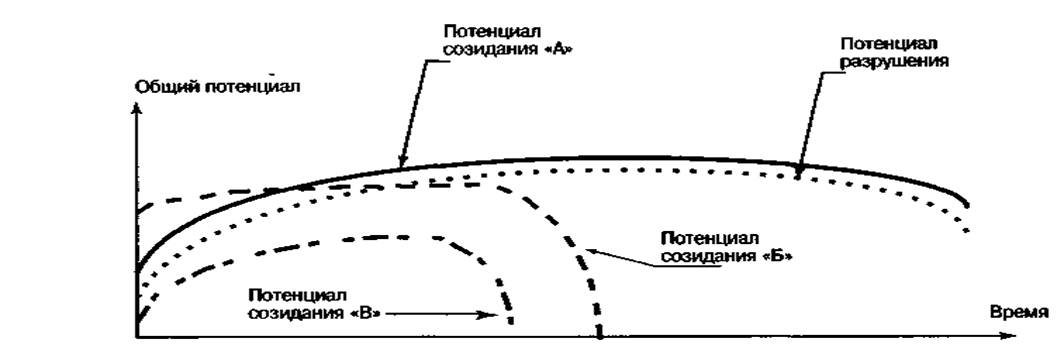

... не знают о законе. Второй вариант. Руководитель знает о законе, а его подчиненные нет. Третий вариант. Руководитель и подчиненные знают о законе», единства анализа и синтеза Список использованной литературы 1. Алиев В.Г., Варфоломеев В.П. Теория организации, курс лекций. «Самоорганизация в природе и обществе», Махачкала ИПЦ ДГУ, 1998. 2. Доблаев В.Л. Теория организаций. — М.: Институт ...
... не тепловая энергия. Вероятно, для выяснения истины необходимо более широкое, чем термодинамическое или статистическое, определение энтропии. Поэтому-то мы рассматриваем этот вопрос здесь, в разделе биологических концепций. Энтропия и эволюция. Вероятно, зарождение проблемы взаимосвязи между энтропией и эволюцией произошло в 1854 г., когда Гельмгольц и Больцман первыми обратили внимание на ...
0 комментариев