Определение величины работы, совершаемой в системе
Если k > 0
Ионное взаимодействие
Гидрофобные взаимодействия
Вращательная диффузия К=0,34 сек
Оба отсека по отдельности должны быть электронейтральными, то есть в каждом отсеке число "+" ионов должно быть равно числу "–" ионов
Связывание проникающей молекулы с определенным участком внутри канала или вблизи него
Эти виды ионной проницаемости обнаруживают самостоятельную изменчивость в зависимости от функционального состояния клетки
Навигация
Гидрофобные взаимодействия
Конспект лекций по биофизике
92536
знаков
0
таблиц
0
изображений
7. Гидрофобные взаимодействия
базируются на силах специфического отталкивания между неполярными атомными группами и молекулами воды. Гидрофобные взаимодействия определяют свойства воды, как конденсированной жидкости. Эти взаимодействия объясняются энтропийным эффектом (реализуется наиболее верояное состояние). Гидрофобное взаимодействие определяет структуру белковых молекул, а также многих надмолекулярных систем.
Особенности строения воды
Вода является средой с большим количеством водородных связей, именно они определяют особые свойства воды:
- ее способность сохранять жидкое состояние в широком диапазоне t0,
- низкую t0 образования льда.
Для льда характерно строго упорядоченное расположение молекул воды, под тетраэдрическими углами. Каждая молекула имеет 4 водородных связи между соседними молекулами. Это рыхлая структура. Когда из льда образуется вода, то эта структура разрушается, в жидкой воде встречаются молекулы с 4,3,2,1 и 0 водородными связями. Водородные связи в жидкой воде характеризуются широким набором углов и длин.
Потенциальная энергия. U вод. связ. жидк. воды – непрерывная функция угла межмолекулярной водородной связи и геометрических характеристик молекулы воды.
Существует несколько моделей структуры воды
1. Разработана Айзенбергом и Кауцманом. Особое внимание обращается на масштаб времени наблюдения за структурой. Удалось обнаружить 3 структуры воды:
1) если сделать снимок с длительностью экспозиции намного меньше, чем период колебаний молекул воды (t<<tкол., t=2*10–13 c), то мы регистрируем М-структуру, которая характеризуется малой упорядоченностью.
2) если продолжить t>>tкол., но намного меньше времени вращения тел диффузии t=10–5 с, то удается пронаблюдать К-структуру. Она характеризуется упорядоченным расположением молекул воды, но случайностью ориентации.
3) при t>>периода вращения диффузии получается Д-структура. Она характеризуется регулярным расположением молекул воды и их правильной, закономерной ориентацией.
2. Кластерная модель Шерага.
Жидкая вода состоит из отдельных молекул и структурно связанных кластеров. Кластеры постоянно распадаются и возникают вновь. Это создает усредненное окружение для каждой отдельно взятой молекулы воды, - слабо учитывает молекулы воды в молекулярных группах.
3. Модель Самойлова
рассматривает структурные изменения воды при различных температурах. Предположим, что во время таяния льда, оторвавшаяся молекула воды заполняет пустоты кристаллической решетки, при этом увеличивается удельный вес. Максимальный удельный вес воды наблюдается при +40С, при более высокой t0 происходит увеличение амплитуды колебаний молекул воды, увеличение занимаемого ею объема и снижению плотности.
Растворимость различных веществ в воде
В воде хорошо растворяются электролиты вследствие высокой диэлектрической проницаемости воды, так же вещества с большим дипольным моментом и вещества, способные образовывать водородные связи с молекулами воды.
Рисунок. Нерастворимые вещества в воде: различные углеводороды, масла, жиры. Это объясняется тем, что контакты между молекулами Н2О–Н2О и С6Н6–О молекулами оказываются более выгодными, чем С6Н6–Н2О. В любой ситуации, когда свободная энергия раствора меньше свободной энергии воды и растворимого вещества, данное вещество хорошо растворяется в воде (и наоборот).
Гибкость полимерных молекул
Молекулы биополимеров сложны и обладают набором свойств.
Инфрмационность полимерных молекул базируется на разнообразии мономерных звеньев. Любая био молекула представляет собой своеобразный био текст и несет в себе некоторый объем информации. Для биофизики важны свойства и информация молекул, для этого информационные молекулы сравнивают с модельными молекулами. К ним относят каучук, он обладает эластчностью (свойство, характерное для биополимеров). Эластичность – это способность полимера испытывать большие упругие деформации (достигающие 100%) при малом модуле упругости. Но в каучуке нет информации, так как он состоит из мономеров, это достоинство используется для сравнения с биополимерами для выяснения биофункции.
Молекулы каучука подчиняются законам Гука
σ = ε(L0–L)/L0,
напряжение σ равно модулю упругости ε, L0 – начальная длина, L – конечная длина. Каучук обладает некоторыми свойствами идеального газа. Энтропийный характер упругости для идеального газа означает, что при движении поршня внутри цилиндра и росте давления в нем, мы переводим его из более вероятного состояния в менее верояное сжатое состояние, понижается энтропия газа. С каучуком происходит тоже самое. Между элементами каучука, связанными в линейную цепь, существуют связи между которыми могут происходить вращения, в результате чего изменяется конформация цепи. В биополимерах так же есть такие связи: С-С, С-N, С-О, вокруг которых могут происходить вращения – молекулы биополимеров обладают конформационной лабильностью. Эти конформационные свойства играют важную роль, так как на них базируются все функционально важные свойства биополимеров.
Клубок, глобула и условия их существованиия
Рисунок. Благодаря вращению вокруг единичных связей, цепочка биополимеров сворачивается самопроизвольно в клубок.
N – количество звеньев
l – средняя длина звена (длины звеньев не равны)
h – расстояние между началом и концом
Нужно найти размер клубка.
h=0, так как конец цепочки может равновероятно находиться в любом месте по отношению к началу, поэтому находят h2 , так как он не равен нулю.
h2=Nl2*(1+cosQ)/(1-cosQ)
рисунок. Q – угол вращения, угол между продолжением и звеном цепи. h2 характеризует размеры полимерного статического клубка. Состояние клубка является наиболее вероятным состоянием биополимера. Ему соответствует максимальная энтропия.
Есть прямые доказательства существования клубка биополимеров. Фотографии. При сворачивании в клубок между атомами возникают взаимодействия двух типов.
1) Взаимодействия ближнего порядка – взаимодействия между соседними полимерными звеньями.
2) Дальние взаимодействия, очень объемные эффекты. Они возникают между атомами, которые в цепочечной структуре биополимера отстоят далеко друг от друга, но вследствие изгибов цепи оказались на небольшом расстоянии. Вследствие объемных эффектов плотность звеньев в пространстве, занятом молекулой биополимера, может изменяться от точки к точке. Существует пространственная корреляция. В состоянии клубка флукитуация (колебания) плотности имеет порядок самой плотности. Однако наличие объемных взаимодействий может привести к такому состоянию в котором флуктуация плотности окажется малой по сравнению с плотностью. Такое состояние носит название глобулы.
Условия существования клубка и глобулы.
Важны заряды, расстояния между мономерами и t0. Увеличение t0 способствует отталкиванию звеньев, снижение приводит к притягиванию. Существуют t0 при которых отталкивание между мономерами полностью компенсируется их взаимным притяжением. Такая t0 соответствует точке Гетта Q (тепла). В Q-точке макромолекула представляет собой клубок с размерами R ≈ lN1/2. При увеличении t0 выше Q-точки возрастают силы отталкивания между мономерам и R > lN1/2 но макромолекула будет в виде клубка. При снижении t0 ниже Q-точки в объемных взаимодействиях будут преобладать силы притяжения между мономерам. Это приведет к конденсации полимерного клубка в плотное слабо флуктуирующее образование, которое называют глобулой, R ≈ lN1/3. Таким образом изменение t0 приводит к изменению размеров макромолекулы, изменению плотности мономеров, и как следствие к изменению энергии взаимодействия и изменению агрегатного состояния.
Свободная энергия взаимодействия звеньев зависит от плотности агрегации этих звеньев.
Рисунок. Вид клубка при нулевой температуре, F- свободная энергия, n – число звеньев. В состоянии клубка молекула имеет min свободной энергии при N ≠ 0. Где F=0 будут осуществляться обратимые переходы между клубком и глобулой. Переходы могут быть двух видов:
1) переходы первого рода: при изменении t0 наблюдается тепловой эффект, S и внутреняя энергия изменяются скачками.
2) фазовый переход второго рода: без тепловых эффектов. Теплоемкость при этом изменяется скачкообразно, S и внутреняя энергия изменяются плавно. В результате удельный V системы не испытывает скачкообразность изменений.
Таким образом вид перехода определяется свойствами макромолекулы. В случае жесткой полимерной цепи переход клубок-глобула осуществляется как фазовый переход первого рода, в случае гибкой цепи – как фазовый переход второго рода.
Рисунок. Графическая зависимость плотности мономерных звеньев от t0. n – плотность мономерных звеньев, 1 жесткая цепь, 2 гибкая цепь. В случае гибкой цепи нет конкретной Q точки, выделяется Q лишь область. В реальных био молекулах гибкость цепи может изменяться в силу различий отдельных участков.
Статистическая картина фазового перехода усложняется в реальных молекулах. Структуры перестройки зависят от физической природы взаимодействий между мономерными звеньями и необязательно усредняются по всему объему, занятому данной молекулой.
Статистическая теория полимерных цепей
СТПЦ берет начало в 50х годах ХХ века из Ленинграда. Основная идея СТПЦ заключается в том, что в полимерной цепи реализуются не любые повороты атомных групп вокруг единичных связей, но существуют лишь определенные поворотные изомеры. Конформацию ротомеров можно установить, если мы знаем химическую структуру цепи.
Рисунок. Этан. Более выгодня транс-конформация, так как вокруг единичной связи вращается молекула и меняется Е потенц.
Рисунок. Графическая зависимость Е потенц. от угла вращения. φ=0 при транс. При поворотах вокруг единичной связи молекула этана преодолевает своеобразный энергетический барьер = 12200 Дж/моль.
Величина энергетического барьера имеет в своей основе энергию дисперсионных сил, если взаимодействующие звенья не полярны; если же они полярны, то кроме дисперсионных сил, свой вклад вносят ориентационные и индукционные силы.
Рисунок. Бутан. СН3–СН2–СН2–СН3 энергетически более выгодна транс-конформация, при которой СН3 группы находятся на max расстоянии друг от друга. Время превращения одного ротомера в другой 10–10 с. Ротомеры нельзя разделить, они непрерывно переходят из одной конформации в другую.
Биофизика клетки. Мембранология.
Все клетки окружены цитоплазматической мембраной, которая представляет собой функциональную структуру, толщиной в несколько молекулярных слоев, которая ограничивает цитоплазму и большинство внутриклеточных структур, а так же образует единую систему канальцев, складок и замкнутых полостей, расположенных внутри клетки. Толщина редко превышает 10 нм, в этой структуре плотно упакованы липиды и белки, поэтому сухой вес мембраны составляет более ½ сухого веса клетки.
В середине XIX века Дюбуа-Реймон впервые сообщил, что между внутренней и внешней поверхностью кожи лягушки имеется разность потенциалов. Моль ввел термин "мембрана" , он изучал цитоплазму клеток растений и выяснил, что она окружена полупроницаемой мембраной. 1877 г. Пфейфер-ботаник, исследуя явление осмоса, пользовался как естественной, так и искусственной мембраной из осадочного ферроцианида. Cu → cходнства между ними → естественная мембрана участвует в явлении осмоса. Позднее стали говорить о генерации биопотенциала мембраны (конец XIX века). 1902 г – Бернштейн – мембранная теория потенциала покоя и потенциала действия → развитие мембранологии. Хаксли, Ходжкин и К0 впервые показала, что потенцилы покоя и действия базируются на избирательной проницаемости мембраны к определенным ионам (К+) – неодинаковое распределение ионов по обе стороны мембраны, в основе чего лежат процессы активного транспорта ионов через мембрану. С участием мембраны связаны: фоторецепция, рецепция, БАВ, передача нервного импульса, синтез ДНК.
Химический состав мембраны
Достаточно высокое содержание липидов, они составляют мембранную матрицу; белки составляют вариабильную часть; углеводы в виде гликопротеидов и гликолипидов. В мембране всегда находится небольшое кол-во воды (важная роль).
Липиды мембраны:
Классификация
I. Классы:
1. Липиды – производные глицерина. Кефалины – фосфодиэтаноламин, Лецитин – фосфатидилхолин.
2. Липиды – производные сфингозина. Сфингомиолин, цереброзиды.
3. Стерины – холестерин, β-ситостерин, эргостерин, зимостерин и т.д.
4. Минорные липиды - β-каротин, витамин К.
II. Группы:
1. Нейтральные липиды – холестерин, триглицериды.
2. Цвиттерионы – 2 заряда диполя – фосфотидилэтаноламин, фосфотидилхолин.
3. Липиды – слабые кислоты, фосфотидилсерин.
4. Липиды – сильные кислоты – фосфотидиловые кислоты и сульфокислоты.
Мембранные белки
С трудом поддаются выделению, многие вообще не выделяются без нарушения структуры. Белки в мембране отличаются большим разнообразием. Большинство белков в мембране находятся в виде клубка, 30% белков могут находится на поверхности мембраны в виде спирали. Существует несколько классификаций мембранных белков:
I. Функциональная классификация
1. ферментативные,
2. транспортные,
3. рецепторные,
4. каналообразующие,
5. воротные,
6. структурные.
II. Классификация по локализации по отношению к липидам.
1. интегральные,
2. периферические.
Интегральные белки погружены в мембрану или пронизывают ее насквозь. Периферические белки на поверхности мембраны и слабо связаны с ней – слабые взаимодействия. На поверхности интегральных белков имеется значительно меньше участков, несущих электрический заряд, чем на поверхности периферических белков.
Вандеркой и Капалди 1972 г. – все аминокислоты делятся на полярные, неполярные и промежуточные. Полярность а-к первой группы приняли за 1, полярность а-к второй группы – за 0, полярность а-к третьей группы – за ½. Изучили состав периферических и интегральных белков: 20 видов мембранных белков: средняя полярность всех белков равна 0,46; интегральные белки имеют полярность от 0,3 до 0,4; периферические белки имеют полярность от 0,41 до 0,53.
Углеводы мембран
В связанном виде не встречаются. В состав мембранных углеводов входят следующие сахара:
- Д-галактоза,
- Д-глюкоза,
- ацетилглюкозамин,
- ацетилгалактозамин,
- Д-фруктоза,
- Д-манноза,
- Д-ксилоза.
Родопсин – гликопротеин оболочки сетчатки, состоит из углеводородной цепочки (4%), связанной с белками М=28000 Да. Гликопротеиды являются рецепторами для гормонов, медиаторов, пептидов и др. Большое кол-во гликопротеидов в вирусных оболочках (до 40% оболочки).
Вода
С ней связаны многие структурно-функциональные свойства мембран, а так же процессы стабилизации и формирования мембран. Вода входит в состав мембран и делится на свободную, связанную и захваченную. Связанная и свободная вода различается по подвижности молекул воды и растворяющей способности. Наименьшей подвижностью и растворяющей способностью обладает внутренняя связанная вода. Она присутствует в липидной зоне мембран в виде отдельных молекул. Основную часть связанной воды представляет вода гидратных оболочек. Эта вода окружает полярные группы белков и липидов, имеет min подвижность и практически не обладает свойствами растворителя. Свободная вода в порах и каналах. По ней могут перемещаться свободные ионы. Она является хорошим растворителем, подвижная и обладает всеми свойствами жидкой воды. Захваченная вода обладает изотопным движением, характерным для жидкой воды, является хорошим растворителем. Она встречается в центральной зоне мембран, между ее липидными слоями, но эта вода пространственно делится как с внеклеточной жидкостью, так и с цитоплазмой. У нее нет возможности свободно с ними обмениваться.
Струкрурная организация мембран
Плохо изучена, но внедряется электронная микроскопия, ЯМР, ЭПР.
Мембраны – 3х слойная структура с наружным и внутренним слоем, тонкие, темные до 2,5 нм, внутренний слой между ними светлый до 3,5 нм. Считается, что основой биомембран в большинстве случаев являются мембранные липиды. 1925 г. Грейбель и Гортер описали свойство фосфолипидов самопроизвольное образованием ими угла биомолекулярного слоя с замкнутой поверхностью в водной среде. Липиды образуют шаровидные образования.
Рисунок. В другой работе показано, что такие образования сохраняют устойчивость, если внешний d этой замкнутой поверхности не меннее 30 нм, так как чем меньше d, тем зазоры между головками липидов больше и вода просачивается внутрь замкнутого образования и нарушает стабильность. Стабильность бислоя определяется заряженными головками липидов.
Ассимметричность – 2ух слоев, они могут состоять из разным липидов.
Эритроцитарная мембрана: во внешнем слое много фосфатидилхолина и сфингомиелина, во внутреннем слое много фосфатидилэтаноламина и фосфатидилсерина. Во многих случаях неполярные хвосты содержат цепочки от 10 до 22 атомов С, между которыми могут быть насыщенные и ненасыщенные связи, это обуславливает ряд свойств мембран. Чем больше ненасыщенных связей, тем ниже t0 замерзания липидного бислоя. Внутренний слой бислоя нерыхлый, он содержит множество холестерина, он заполняет пространство между неполярными хвостами, влияет на t0 замерзания бислоя: чем больше холестерина, тем ниже t0 кристаллизации. Холестерин участвует в стабилизации мембран и будет влиять на проницаемость мембран, чем его больше, тем ниже проницаемость мембраны.
Мембранные липиды обладают динамическими свойствами:
- способность липидной молекулы к латеральной диффузии, коэффициент латеральной диффузии равен 3,25*10–8 см/сек. Коэф отражает способность перемещения липидной молекулы вдоль мембраны,
- вращательная диффузия, К=10–9 сек,
- flip-flop переход, липидные молекулы пересекают мембрану, переходя из одного слоя в другой. К=10–3 сек средняя величина, показывающая число переходов – 1 переход в 1000 сек.
Организация мембранных белков
Большая часть мембранных белков находится в виде клубка (≥70%), основная часть может разворачиваться на поверхности липидного бислоя вследствие электростатического взаимодействия с липидными головками. В этом случае белки будут расположены на поверхности липидов в виде спирали.
Родоспин, М=28000 Да, форма сферы, d=4 нм, мелкая молекула.
Динамические свойства белков.
1. Латеральная диффузия. все значения для белков с М=100000, К=3*10–10 см/сек. Но белки могут объединяться в кластеры, которые мало подвижны.
Похожие работы
... Во времена, например, Платона. Они были одни, а на современном этапе они другие. По мнению Карпекова К.С.(2).(который по моему мнению наиболее полно обозначил современную концепцию происхождения объектов Вселенной.). В.И. Вернадский впервые поставил вопрос о рациональном контроле над окружающей средой. Ноосфера Вернадского – это сфера материального, в котором в качестве управляющего, регулирующего ...
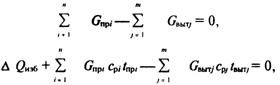
... или технологических процессов; – при выборе технического решения обеспечить малоотходность производства и максимальную эффективность использования энергоресурсов. Задачи специалиста в области безопасности жизнедеятельности сводятся к следующему; – контроль и поддержание допустимых условий (параметры микроклимата, освещение и др.) жизнедеятельности человека в техносфере; – идентификация ...
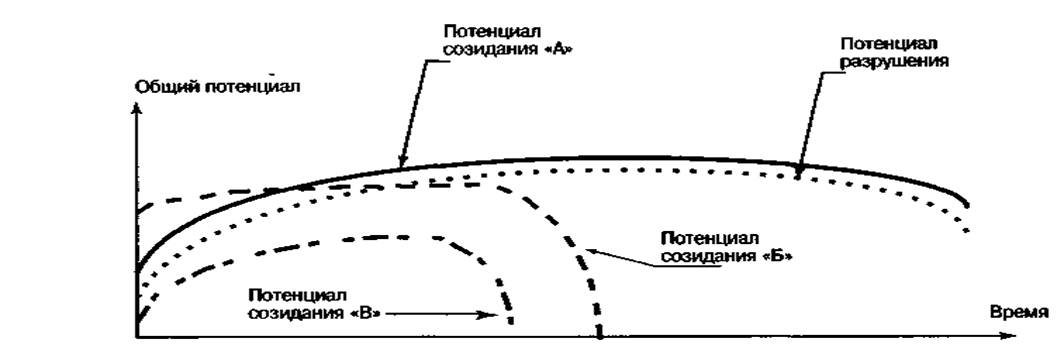

... не знают о законе. Второй вариант. Руководитель знает о законе, а его подчиненные нет. Третий вариант. Руководитель и подчиненные знают о законе», единства анализа и синтеза Список использованной литературы 1. Алиев В.Г., Варфоломеев В.П. Теория организации, курс лекций. «Самоорганизация в природе и обществе», Махачкала ИПЦ ДГУ, 1998. 2. Доблаев В.Л. Теория организаций. — М.: Институт ...
... не тепловая энергия. Вероятно, для выяснения истины необходимо более широкое, чем термодинамическое или статистическое, определение энтропии. Поэтому-то мы рассматриваем этот вопрос здесь, в разделе биологических концепций. Энтропия и эволюция. Вероятно, зарождение проблемы взаимосвязи между энтропией и эволюцией произошло в 1854 г., когда Гельмгольц и Больцман первыми обратили внимание на ...
0 комментариев