Навигация
Регистрация сигнальных молекул
41813
знаков
6
таблиц
0
изображений
содержание
| стр. | |
| Список сокращений | |
| введение | |
| 1. обзор литературы | |
| 1.1. Использование Ti-плазмид агробактерий в генетической инженерии | |
| 1.1.1. Краткая характеристика Agrobacterium tumefaciens | |
| 1.1.2. Cоздание векторов на основе Ti-плазмид | |
| 1.1.3. Процессинг тДНК в бактериальной клетке и ее перенос в клетки растений | |
| 1.1.4. Разработка система трансформаций растений с помощью Agrobacterium tumefaciens | |
| 1.1.5. Проблема сохранения чужеродных генов, перенесенных в растение | |
| 1.1.6. Анализ экспрессии чужеродных генов в трансформированных растениях | |
| 1.2. Использование метода генетической инженерии для трансформации однодольных растений | |
| 1.2.1. Краткие характеристики ряски | |
| 1.3. Сигнальные молекулы, их способность индуцировать процессинг тДНК | |
| 2. Материалы и методы | |
| 2.1. Материалы | |
| 2.1.1. Оборудование | |
| 2.1.2. Бактериальные штаммы и плазмиды | |
| 2.1.3. Растения | |
| 2.1.4. Среды микробиологические для культивирования растений | |
| 2.1.5. Другие растворы | |
| 2.1.6. Ферменты, используемые в генной инженерии | |
| 2.1.7. Антибиотики | |
| 2.2. Методы | |
| 2.2.1. Инкубация Agrobacterium tumefaciens с экссудатами тканей растений | |
| 2.2.2. Выделение тотальной ДНК Agrobacterium tumefaciens | |
| 2.2.3. Блод-гибридизация тДНК по Саузерну | |
| 2.2.4. Трансформация клеток Esherichia coli | |
| 3. результаты | |
| 4. обсуждение | |
| 5. выводы | |
| литературы | |
| приложение |
СПИСОК СОкРАЩЕНИЙ
НУК – нафтиуксусная кислота
БАП – бензиламинопурин
MS –
LB –
тДНК –
Ар – ампицилин
Cs – цефотоксин
ВВЕДЕНИЕ
Актуальность работы:
Свойство бактерий вида Agrobacterium tumefaciens вызывать у растений корончато-галловую болезнь связано с присутствием в их клетках крупных (95-156 мДа) конъюгированных Ti-плазмид (от англ. tumor-inducing – вызывающий опухоль). В процессе идентифицирования растений часть генетического материала Ti-плазмид – тДНК (от англ. transferred DNA – передаваемая) перемещается в растительные клетки и интегрируется в хромосомы, оставаясь частью наследственного материала. Гены тДНК экспрессируются в трансформарованных растительных клетках, нарушают их фитогормональный баланс и определяют синтез специфических ------- соединений.
Таким образом, агробактерии являются природными "генными инженерами", осуществляющий поразительный по филогенетической дальности перенос генетической информации. На основе агробактерий сконструированы эффективные векторные системы для генетической инженерии растений.
Агробактириальная трансформация происходит в результате сложного процесса взаимодействия между бактериальными и растительными клетками. В этом процессе одной из решающих стадий является рецепция агробактериями особых сигнальных молекул, присутствующих в экссудатах поврежденных тканей растений. Сигнальные молекулы индуцируют экспрессию генов области vir (агробактериальных Ti-плазмид), контролирующих вырезание тДНК и ее перенос в клетки растений. Агробактерильная трансформация наблюдается у широкого круга голосеменных и двудольных растений, однако, она отмечена лишь у весьма незначительного числа однодольных растений. Одной из причин ограничений агробактериальной трансформации однодольных считается присутствие в их клетках сигнальных молекул, индуцирующих процессинг и перенос тДНК.
Цель работы:
В настоящее время имеются противоречивые данные относительно наличия таких сигнальных молекул у однодольных растений. В связи с этим, целью данной работы был анализ ряски на присутствие у них сигнальных молекул, индуцирующих процессинг тДНК.
1. ОБЗОР ЛИТЕРАТУРЫ
1.1. Использование Ti-плазмид агробактерий в генетической инженерии растений
1.1.1. Краткая характеристика Agrobacterium tumefaciens
За последнее десятилетие в области генетической инженерии растений достигнуты значительные успехи. Были разработаны разнообразные методы генетической трансформации и, в настоящее время осуществлена экспрессия чужеродных генов в растениях многих видов. Наиболее важным для развития генетической инженерии растений было открытие молекулярных основ опухолевых образований с помощью Agrobacterium tumefaciens [7].
Вирулентные штаммы Agrobacterium tumefaciens (сем. Rirobiaceae) характеризуются присутствием в клетках большой плазмиды, так называемой Ti-плазмиды, весом более 150 т.п.н. (см. Приложение) [9].
Агробактерии вызывают опухолевый рост у многих двудольных и голосеменных, а так же у некоторых однодольных растений [2,3]. Для инфицирования in vivo необходимо повреждение тканей растения [12].
После прикрепления к клеточной стенке растительной клетки агробактерии переносят часть Ti-плазмиды (так называемой тДНК) в ядро, где происходит ее стабильная интеграция в хромосому растения.
Доказано, что функция распознавания клеток и прикрепления к ним, а так же вырезание, перенос и, возможно, интеграция тДНК в растительный геном кодируется двумя хромосомными генами – Chva и Chvb [13] и рядом генов vir-области, находящихся на Ti-плазмиде [14].
После переноса в ядро растительной клетки, тДНК может интегрировать в геном в виде одной или нескольких копий [15]. Встроенная тДНК имеет свойства, характерные для ДНК эукариот, что показано в экспериментах по гиперчувствительности к ДНК-азе I (Schafer, 1984). В зависимости от типа Ti-плазмиды, в тДНК находится от семи до тринадцати генов, ответственных за опухолевый фенотип. Гены 1 и 2 кодируют ферменты, участвующие в синтезе ауксина, индолилуксусной кислоты, в то время, как ген 4 кодирует изопентенилтрансферазу, синтезирующую цитокинин изопентенила денозин 5'-монофосфат [16].
Одновременная транскрипция генов 1,2 и 4 приводит к повышению уровня фитогормонов внутри трансформированных клеток. Результатом этого является повышение митотической активности и образование опухоли. Другие гены тДНК кодируют синтез так называемых опинов, из которых наиболее изучены нопапин и октопин. Опины представляет собой производное аминокислот и сахаров, которые служат источником питания для агробактерий [14]. В целом, образование корончатого галла представляет собой хорошо охарактеризованный пример генетической инженерии растений в природе.
Мутации в вирулентных генах агробактерий. Наличие Ti-плазмид в клетках агробактерий является абсолютно необходимым условием патогенности микроорганизма. Излеченные от Ti-плазмид штаммы агробактерий авирулентны. На вирулентность Agrobacterium tumefaciens оказывают влияние различные мутации, картируемые как на опухолевых плазмидах, так и на хромосомах. Ранние этапы взаимодействия агробактерий с растениями, так же как хемотаксис, прикрепление к поверхности растительной клетки и специфическое связывание в центрах инфекции контролируются генами, имеющими хромосомную локализацию. В хромосоме расположены некоторые гены, регулирующие экспрессию vir-генов Ti-плазмид [19]. Присоединение агробактерий к клеткам растения является одним из первых этапов, определяющих эффективное взаимодействие. Этот этап у Agrobacterium tumefaciens контролируют два связанных между собой хромосомных локуса Chva и Chvb, размерами 1,5 kb и 5kb, соответственно [Дуглас и др, 1985]. Гены этих локусов экспрессируются конститутивно. В результате транспозонного мутагенеза этих областей получают авирулентные или дефекнтые по прикреплению агробактерии. Мутации в этих локусах сильно понижают или ингибируют вирулентность бактерии, но не для всех хозяев. Локус Chvb определяет синтез нейтрального циклического b-D-гликана, который трансформируется в периплазматическое пространство клетки с помощью продукта гена Chva. Роль нейтрального b-D-гликана в инфекционном процессе еще точно не установлена. Помимо циклического b-D-гликана в прикреплении патогенных агробактерий к растительным клеткам принимают участие и другие полисахариды, в частности внеклеточные экзополисахариды.
Организация vir-генов Ti-плазмид. Вирулентные гены агробактерий на Ti- и Ri-плазмидах кластеризованы в области vir разметом около 30-35 kb, проявляющей в этих плазмидах значительную гомологию ДНК. Выявлена также гомология vir-генов Ti-плазмид Agrobacterium tumefaciens с tra-генами конъюгитивных плазмид. В ----- Ti-плазмидах в области vir локализовано шесть различных групп комплементации A, B, C, D, E и G, организованных в единый регулон [Stachel Nester, 1986]. Октопиновая Ti-плазмида Arh 5 имеет дополнительный локус vir F, расположенный справа от локуса vir E [Kooykaas et al., 1984]. Мутации в генах и --------- vir A, vir G, vir B и vir D придают агробактериям авирулентный фенотип, в отличие от большинства хромосомных мутаций, имеющих круг трансформируемых растений-хозяев.
Продукты генов vir-области контролируют процессинг тДНК в бактериальной клетке, ее перенос в растительную клетку и интеграцию в ядерный геном растения, причем эти процессы гены vir могут определять не только в цис, но и в транс положении по отношению к тДНК (то есть находясь в разных репликонах). Исходя из этого свойства области vir, сконструированы и успешно используются в практике удобные бинарные векторы для генетической инженерии растений [Дрейпер с соавт, 1991]. Для процессов "вырезания" тДНК из плазмиды (точнее, высвобождения в процессе репликативного синтеза) и ее переноса в растение необходимо фланкирование этой области особыми границами: несовершенными прямыми повторяющимися последовательностями ДНК размером 24 ---- , проявляющими значительную гомологию у всех изученных Ti- и Ri-плазмид. Границы тДНК гомологичны области oriT конъюгативных плазмид. В этой области сайт-специфические эндонуклеазы производят одноцепочечный разрыв, служащий началом репликации по типу разматывающегося рулона, происходящей в процессе транспорта плазмиды. Репликация обеспечивает сохранение плазмиды в материнской клетке и появление ее копии в дочерней.
Для нормального процессинга тДНК и ее переноса в растительную клетку особенно важна ее правая граница, которая одна может определять полярность переноса тДНК. Удаление правой границы из Ti-плазмид делает агробактерии полностью авирулентными. Замена ее на искусственно синтезированную, так же как и на левую, восстанавливает вирулентность микроорганизма.
На процессинг тДНК в клетках бактерий влияют мутации в генах vir D, vir C и vir E – оперонов, на транспорт т-комплекса в растительную клетку – мутации в генах vir B и vir D – оперонов.
1.1.2. Создание векторов на основе Ti-плазмид
В начале восьмидесятых годов были сделаны первые попытки перенести чужеродные последовательности ДНК в растительные клетки либо с помощью транспозонного мутагенеза [Uernals-teens et al., 1980], либо путем сайт-специфической миграции генов в тДНК и последующей двойной рекомбинацией с Ti-плазмидой дикого типа [Matrke et al., 1981; leemans et al., 1981]. Однако, эти ранние эксперименты, основанные на двойной рекомбинации, занимали много времени, были довольно сложны и трансформации проходили с очень низкой частотой. Необходимо было разработать более эффективные векторы, чтобы облегчить генетические манипуляции с бактериями и позволить селекцию и регенерацию трансформатов.
Сейчас используют две принципиально разные системы для введения чужеродных генов в растения с помощью Ti-плазмид:
1. ---- векторы
2. бинарные векторы.
В основе создания -------- векторов лежит тот факт, что гены тДНК не ------- для растительных клеток, и любая последовтельность ДНК, встроенная между границами тДНК, может интегрировать в хромосому растительной клетки и нормально там экспрессироваться [Zambryski et al., 1983]. В -------- векторых системах тДНК можно заменить, например, на последовательность pBR322, а чужеродную ДНК, которую предполагается перенести в растения, нужно проклонировать в этом же векторе.
Затем путем гомологичной рекомбинации эта чужеродная ДНК может быть перенесена на Ti-плазмиду реципиентного штамма агробактерии (рис. 2). Одним из первых таких векторов на основании Ti-плазмид авляется pGV3850 [Zambryski et al., 1983]. В нем все гены, ответственные за синтез фитогормонов, были заменены на последовательность pBR322.
ДНК pBR322 обеспечивала гомологию для ------- области тДНК pGV3850 с любыми производыми pBR, несущими клонированный ген.
Гены, кодирующие различные маркерные белки для быстрого отбора трансгенных растений, были встроены в pGV3850 [De Blocle, 1984]. Была разработана система трехродительного скрещивания для переноса любых производных pBR322 из E. coli в A. tumefaciens pGV3850 [Van Haute et al., 1983]. В настоящее время сконструированы и успешно используются и другие --- ---------- векторы на основе Ti-плазмид [Royers et al., 1988].
Рис. 2.
Схемы -------- (А) и бинарной (Б) векторных систем. vir – область вирулентности. HOM – области гомологии, в пределах которых может происходить рекомбинация, приводящая к образованию коинтегратов. LB и RB – левая и правая границы тДНК. MCS - ----- сайт для клонирования. РТМ – маркет трансформации для растений. RES – маркер устойчивости к антибиотику для бактерий. OriT – начало переноса и bom-сайт для мобилизации векторов при конъюгации. Col E1 – начало репликации из плазмиды Col E1. RK2 – начало репликации из плазмиды широкого круга хозяев RK2.
Система бинарных векторов основана на том, что область тДНК и гены vir могут распологаться на разных плазмидах [Hockemu et al., 1983]. В таких системах обычно присутствуют два элемента:
1) Ti-плазмида-помощник, в которой тДНК польностью делетирована. Эта плазмида несет в своем составе гены vir, действующие in trans.
2) Плазмида широкого круга хозяев, имеющая сайты для клонирования и маркерные гены для селекции растений, ограниченные правой и левой фланкирующими последовательностями тДНК [An et al., 1988] рис
Обе описанные выше системы векторов предполагают --------- этапах сборку нужных конструкций в промежуточных векторах, например в pAP2034 [Veltena, Sehell., 1987] или pRT103 [Topter et al., 1983] а затем перенос из в готовом виде в рецепиентные штаммы агробактерий.
Похожие работы
... очередь, активизирует ферменты — киназы подобно тому, как цифровая информация наносится на поверхность компакт-диска. Участки мозга человека, отвечающие за различные виды забывчивости: „выветривание“ воспоминаний со временем, рассеянность, заторможенность (например, человек забыл слово, вертящееся на языке), нарушение причинно-следственных связей в воспоминаниях, придуманные воспоминания ...
... Во-вторых, в настоящее время известно достаточное количество соединений, нерастворимых в неполярных растворителях или же, наоборот, хорошо растворимых в воде, которые, тем не менее, относят к липидам. В современной органической химии определение термина «липиды» основано на биосинтетическом родстве данных соединений — к липидам относят жирные кислоты и их производные [1]. В то же время в биохимии ...


... . Очевидно, что научное наследие, оставляемое нам И. И. Мечниковым, и в будущем будет определять основные направления учения о фагоцитах. Макрофаги перитонеального экссудата как модель фагоцитоза и нарушений фагоцитарной активности. В организме человека фагоцитирующую функцию выполняют несколько типов клеток. Прежде всего, это те клетки, которые осуществляют защиту при каких-либо инфекциях и ...
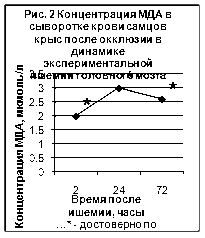

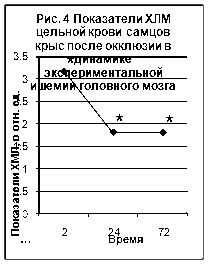
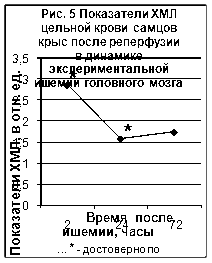
... ТБК-активных продуктов – малонового диальдегида (МДА), показателям люминолзависимой хемилюминесценции. 3.2. Исследование активности хемилюминесценции цельной крови и концентрации МДА в сыворотке крови при экспериментальной ишемии головного мозга Центральное место в изучении ишемического инсульта занимают экспериментальные модели на животных [65]. Именно по результатам эксперимента ...
0 комментариев