Навигация
Выделение тотальной ДНК Agrobacterium tumefaciens
41813
знаков
6
таблиц
0
изображений
2.2.2. Выделение тотальной ДНК Agrobacterium tumefaciens
ДНК выделяли по модифицированному методу Draper с соавт.Через 48 часов совместного культивирования агробактерий с тканями растений из пробирок удаляли растительную ткань, бактерии собирали, центруфугировали при 9000 g. Осадок, полученый из 10 мл инкубационной среды лизировали в течение 45 мин при 370 С в 500 мкл ТЕ буфера (10 мМ трисHCl (pH 8,0), 1 мМ ЭДТА), содержащего 1% сарколизата Na и 1,5 мг/мл проназы. Лизат дважды экстрагировали фенолом и дважды хлороформом. ДНК осаждали спиртом и растворяли в ТЕ–буфере.
2.2.3. Блод-гибридизация тДНК по Саузерну
В целях дополнительной проверки наличия индукции процессинга и присутствия в пробах pBR322 татальную ДНК агробактерий в количестве 5 мкг наносили в лунки 0,8% агарозного геля и проводили электрофорез при 25–100 В в течение 2 часов в буфере, содержащем: 40 мМ трис–ацетат (рН 8,0), 1 мМ ЭДТА и 5 мкг/мл бромистого этидия. В качестве маркеров молекулярного веса использовали ДНК фага l, гидролизованную рестриктазной эндонуклеазой Hind III. Блод–гибридизацию проводили на ––––– фильтрах. В качестве зонда использвали ДНК плазмиды pBR322, меченную 32 –– с помощью ДНК–полимеразной системы.
2.2.4. Трансформация клеток Esherichia coli
Штаммы E. сoli HB101 выращивали в жидкой среде LB при 370 С до концентрации 5*107 клеток/мл (А600 = 0,45). Для количественного изучения процессинга тДНК использовали комплементарные клетки бесплазмидного штамма E. сoli HB101, которые трансформировали тотальной ДНК агробактерий, которая была выделена (М-2), претерпевших различную переработку. Трансформанты каждого варианта высевали на три чашки Петри с агаризированной средой LB, содержащей ампицилин (50 мкг/мл). Количество колоний подсчитывали. В контрольных экспериментах клетки трансформировали плазмидой pBR322, и другой контроль не был подвергнут трансформации и колоний обнаружено не было.
3. результаты
Для обнаружения процессинга агробактериальной тДНК, индуцированного экссудатами анализированных растений, в работе применяли метод "спасения плазмиды", предложенный Koukolikova-Nikola с соавт. Этот метод основан на использовании модифицированной Ti-плазмиды pGV385 [11], содержащей между правой и левой границами значительно делетированной исходной тДНК бактериальной плазмиды вектор pBR322. Таким образом, индуцирование процессинга тДНК в модифицированной Ti-плазмиде pGV3850 можно прослеживать по вырезанию из нее низкомолекулярного фрагмента ДНК, в состав которого входит плазмида pBR322. Этот процесс можно тестировать различными методами. Один из них – это трансформация клеток E. сoli тотальной агробактериальной ДНК и отбор трансформантов по селективным маркерным признакам плазмиды pBR322 – устойчивой к антибиотикам. Другим методом может быть блод-гибридизация – анализ тДНК с помощью плазмиды pBR322 в качестве гибридизационного зонда. Оба этих метода были использованы в настоящей работе. Сравнили эффективность индукции процессинга экссудатами различных растений проведено по отношению к активности 100 мкМ ацетосирингона. Известно, что это токсифенольное соединение, выделяемое некоторыми двудольными растениями, принадлежит к сигнальным молекулам, индуцирующим вырезание и перенос агробактериальной тДНК.
Результаты трансформации клеток E. сoli НВ101 тДНК подвергнутых воздействию экссудатов листьев различных растений представлены в таблице.
Таблица 3.1.
| Факторы индукции | Количество трансформированных E. Coli |
| L. perpusilla (ряска) L. perpusilla (ряска) Ацетосирингон (контроль) Лен долгунец Nicotiana tabacum (табак) Без ацетосирингона и трансформации | 25 колоний 30 колоний 35 колоний 40 колоний 30 колоний – |
Экссудаты листьев ряски индуцировали вырезание тДНК, что является доказательством присутствия в их составе сигнальных соединений, специфических для индукции транскрипции генов vir агробактериальной Ti-плазмиды.
Прямой блод-гибридизационный анализ тДНК агробактерий свидетельствует о присутствии у ряски сигнальных молекул, индуцирующих образование тДНК [12].
4. обсуждение
Использованный подход позволяет регистрировать присутствие сигнальных молекул в экссудатах различных тканей растений по одному из важнейших результатов индукции генов области vir, а именно по вырезанию тДНК из агробактериальной Ti-плазмиды. В настоящей работе мы применили два метода детекции процессинга тДНК модифицированной агробактериальной Ti-плазмиды. PGV3850 [11].
Полученные результаты позволяют расширить список однодольных растений (на одно растение), синтезирующих сигнальные молекулы, индуцирующие процессинг агробактериальной тДНК. Примененный метод "спасения плазмиды" позволяет выявлять появление циклических форм модифицированной тДНК pBR322 и ее линейных форм [20].
Рассмотренный в работе метод позволяет учитывать присутствие в экссудатах растений веществ с бактериостатической активностью. Вещества такой природы могут существенно влиять на эффективность трансформации растений агробактериями.
Мы не анализировали химическую природу сигнальных молекул ряски. Не исключено, что сигнальные молекулы могут отличаться от сигнальных фенольных соединений двудольных растений. Специфический процесс взаимодействия между агробактериями и покрытосеменными растениями существовал, по-видимому, еще до их дивергенции на двудольные и однодольные [15].
Штаммы агробактерий обладают многими особенностями и именно рецепторы (продукты гена vir A), различающиеся по способности узнавать сигнальные молекулы различной природы.
Следует отметить, что растительные вещества с сигнальной функцией для агробактерий не является уникальным примером существования соединений, выполняющих эту важную роль во взаимодействии микроорганизмов с растенями [22].
Молекулярные сигналы растений играют важную роль в установлении паразитических и симбиотических взаимоотношений между бактериями и растениями, определяя при этом круг растений для определенного микроорганизма. Всесторонние структурно-функциональные исследования специфических молекулярных сигналов во взаимодействии между микроорганизмами и растениями должны ответить на многочисленные вопросы о молекулярно-генетических механизмах этих взаимоотношений и способствовать развитию эффективных методов генетической инженерии важнейших сельскохозяйственных и экологически-индикаторных культурах растений (как ряска).
Список литературы
| 1. 2. 3. 4. 5 6. 7. 8. 9. 10. | Анализ генома. Методы. Под ред. Н. Дейвиса. – М.: Мир, 1990. – 247 с. Великов В.А., Бурьянов Я.И. Образование делеционных производных Ti-плазмиды pGV3850 при конъюгационном переносе из Agrobacterium tumefaciens в Escherichia coli. // Генетика.1998. №8. т. 34. с. 1056-1062. Великов В.А., Бурьянов Я.И. Изучение конъюгационного транспорта Ti-плазмиды pGV3850 из Agrobacterium tumefaciens в Escherichia coli. // Тезисы докладов III Пущинской конференции молодых ученых. Пущино, 27-30 апреля 1998 г, с. 49. Генная инженерия растений. Лабораторное руководство. – М.: Мир, 1991. – 408 с. Клонирование ДНК. Методы. Под ред. Д. Гловера. – М.: Мир, 1988. – 538 с. Малиатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. Методы генетической инженерии. – М.:Мир, 1984. – 463 с. Методы молекулярной генетики и генной инженерии. Под ред. А.И. ----. – Новосибирск: Наука, 1990. – 248 с. Мобильность генома растений. Под ред. Б. Хол и Е.С. Делинс. – М.: Агропромиздат, 1990. – 272 с. Плазмиды. Методы. Под ред. Д. Хорда. – М.: Агропромиздат, 1990. – 272 с. Пехов А.П. Основы плазмидологии.- М.: 1996. – 231 с. Сельскохозяйственная биотехнология. Векторные системы молекулярного клонирования. Под ред. Р.Л. Подреперс и Д.Т. Денхардт. – М.: Агропромиздат, 1991. – 535 с. Солова Г.К., Кривопалов Ю.В., Великов В.А., Чумаков М.И. Прикрепление Agrobacterium к корням пшеницы. // Микробиология. 1995. т. 64. №4, с. 526-530. Захарченко Н.С., Комиви М.А., Бурьянов Я.И. Индуцирование процессинга тДНК. // Физиология растений. 1999. т. 46. № 2, - с. 282-291. Baker R.F., Idler I.B. and al. Nucleotide sequence of the tDNA region from Agrobacterium tumefaciens Ti plasmid pTi / 5955. Plant Mol. Biol., 1983, V.2, pp. 335-350. Douglas C.J and al. Identification and genetic analisys of an Agrobacterium tumefaciens chromosomal virulence rigion. J. Bacteriol., 1985, v. 161, pp. 850-860. Holster S.M., Villaroel and al. An analisys of the boundaries of the octopine TL – DNA intumors induced by Agrobacterium tumefacies. Mol. Gen. Genet., 1983, v. 190, pp. 35-38. Howord E.A., Zupan J.R. and al. The vir Dr protein of A. Tumefaciens contaen sal-terminal bipartite nuclear localization sygnal: implication for nuclear uptake of DNA in plant cells. Cell, 1992, v. 68, pp. 109-119. Janssens A., Engler., Zambryski P. The nopaline c58 tDNA region is teansribed in Agrobacterium tumefaciens., Mol. Gen. Genet., 1984, v. 195, pp. 341-350. Melchers L.S., Hooykaas P.J.J., virulence in Agrobacterium tumefaciens. Oxford Surv. Plant Mol. Cell Biol., 1987. № 4. pp. 167-170. Stachel S.E., Nester E.W. The genetic and transcriptional organization of the vir regulon of the A6 Ti plasmid of Agrobacterium tumefaciens. EMBO J., v. 5, pp. 1145-1454. Zambryski P., Joos N., Genetello G., Leemans. Ti plasmid veefof for the introduction of DNA in to plant cells without alteration of their normal regeneration Capacity. EMBO J., 1983. v. 2, pp. 2143-2150. |
Похожие работы
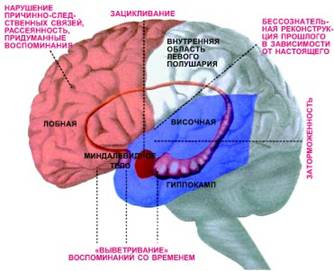
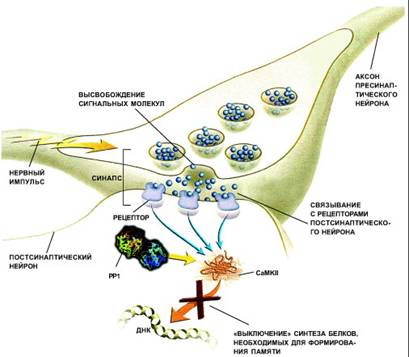
... очередь, активизирует ферменты — киназы подобно тому, как цифровая информация наносится на поверхность компакт-диска. Участки мозга человека, отвечающие за различные виды забывчивости: „выветривание“ воспоминаний со временем, рассеянность, заторможенность (например, человек забыл слово, вертящееся на языке), нарушение причинно-следственных связей в воспоминаниях, придуманные воспоминания ...
... Во-вторых, в настоящее время известно достаточное количество соединений, нерастворимых в неполярных растворителях или же, наоборот, хорошо растворимых в воде, которые, тем не менее, относят к липидам. В современной органической химии определение термина «липиды» основано на биосинтетическом родстве данных соединений — к липидам относят жирные кислоты и их производные [1]. В то же время в биохимии ...


... . Очевидно, что научное наследие, оставляемое нам И. И. Мечниковым, и в будущем будет определять основные направления учения о фагоцитах. Макрофаги перитонеального экссудата как модель фагоцитоза и нарушений фагоцитарной активности. В организме человека фагоцитирующую функцию выполняют несколько типов клеток. Прежде всего, это те клетки, которые осуществляют защиту при каких-либо инфекциях и ...
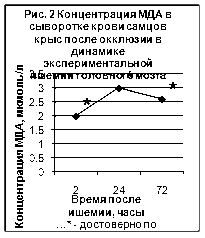

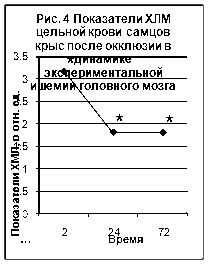
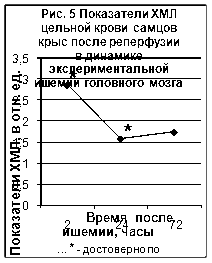
... ТБК-активных продуктов – малонового диальдегида (МДА), показателям люминолзависимой хемилюминесценции. 3.2. Исследование активности хемилюминесценции цельной крови и концентрации МДА в сыворотке крови при экспериментальной ишемии головного мозга Центральное место в изучении ишемического инсульта занимают экспериментальные модели на животных [65]. Именно по результатам эксперимента ...
0 комментариев