Структура молекулы тайтина и его функции
Белки семейства тайтина в норме, при адаптации и патологии
История изучения амилоидозов
Изучение амилоидных фибрилл in vitro
Выделение тайтина из скелетных мышц
Метод флуоресцентного анализа
Изучение вторичной структуры тайтина и белков его семейства до и после образования амилоидных фибрилл
Образование амилоидных фибрилл С-белком миокарда человека при ДКМП и С-белком миокарда кролика
Навигация
Структура молекулы тайтина и его функции
Исследование соотношения в мышцах С- и Х-белков в норме и при патологии
86165
знаков
3
таблицы
7
изображений
1.1 Структура молекулы тайтина и его функции
Тайтин (= коннектин, титин) – гигантский эластичный белок скелетных и сердечных мышц млекопитающих (Maruyama et al., 1977; Wang et al., 1979) с молекулярным весом более 3 МДа (Labeit & Kolmerer, 1995). Он был открыт независимо двумя группами исследователей методом ДСН-гель-электрофореза (Wang et al., 1979; Maruyama et al., 1981). Тайтин является третьим по количеству (после актина и миозина) компонентом саркомера поперечно-полосатых мышц позвоночных. Молекулы тайтина длиной около 1 мкм и шириной 3–4 нм (Trinick et al., 1984; Itoh et al., 1986; Nave et al., 1989; Soteriou et al., 1993; Suzuki et al., 1994; Houmeida et al., 1995; Tskhovrebova & Trinick, 1997) перекрывают половину саркомера от М-линии до Z-линии (рис. 1), простираясь через I- и А-диски саркомера, формируя третью филаментную систему в миофибриллах (Fürst et al., 1988). NH2-концы молекулы тайтина из смежных саркомеров перекрываются в Z-линии, а СООН-концы перекрываются в М-линии. На каждую половину саркомера приходится по шесть молекул тайтина, которые объединены в А-диске саркомера (Trinick 1981; Liversage et al., 2001) и, предположительно, разделяются на пары или на индивидуальные молекулы в I-диске саркомера (Funatsu et al., 1993).
Исследования последних лет показали, что этот гигантский полипептид имеет различное строение в разных зонах саркомера, что вносит свой вклад в их архитектуру и функционирование. Бóльшая часть (~90%) молекулы тайтина состоит из повторяющихся иммуноглобулин-С2-подобных (IgС2) и фибронектин-3-подобных (FnIII) доменов (Labeit & Kolmerer, 1995) с β-складчатой структурой, содержащих около ста аминокислотных остатков (Improta et al., 1996; Muhle-Goll et al., 1998) (рис. 2). Кроме этих доменов тайтин содержит уникальные последовательности, расположенные по всей длине его молекулы: киназный домен вблизи М-линии саркомера; два участка связывания с кальпаиновой протеазой (р94) в I-диске и М-линии саркомера, а также эластичные N2B, N2A и PEVK элементы в I-диске саркомера. В концевых областях молекулы тайтина расположены фосфорилируемые участки.
1.2 Структура и функции молекул С-белка, Х-белка и Н-белка
Предположение о присутствии белков немиозиновой природы в структуре толстых, или миозин-содержащих нитей на основании электронно-микроскопических и рентгеновских исследований высказывались давно (Draper & Hodge 1949; Franzini-Armstrong & Porter 1964; Pepe 1967; Hanson et al., 1971), однако до начала 70-х годов природа этих белков была неизвестна. С-белок, Х-белок и Н-белок были обнаружены с помощью ДСН-гель-электрофореза как минорные примеси в препаратах миозина скелетных мышц кролика (Starr & Offer, 1971). В настоящее время эти белки охарактеризованы, и их количество в саркомере составляет около 5% от массы миозина.
С-белок впервые был выделен из скелетных мышц кролика (Offer et al., 1973). Его количество составляет около 2% от массы миофибрилл и около 4% от массы толстой нити.
Показана способность сердечного С-белка фосфорилироваться разными протеинкиназами в условиях in vivo и in vitro (Jeacocre & England, 1980; Hartzell & Titus, 1982; Hartzell & Glass, 1984; Hartzell, 1985; Lim et al., 1985; Gautel et al., 1995; Mohamed et al., 1998). Молекулярный вес С-белка скелетных мышц по данным ДСН-гель-электрофореза 135 кДа (Yamamoto & Moos, 1983; Starr & Offer, 1983), а по данным анализа нуклеотидной последовательности генов, кодирующих этот белок, составляет 128.1 кДа (Weber et al., 1993). Аминокислотный состав белка характеризуется довольно высоким содержанием пролина, что коррелирует с низким содержанием α-спирали в его молекуле. Методом седиментации показано, что С-белок связывается с высоким сродством с миозином (Callaway & Bechtel, 1981; Yamamoto & Moos, 1983). Электронно-микроскопические исследования подтвердили это связывание для С-белка: он декорирует реконструируемые миозиновые нити и паракристаллы легкого меромиозина (ЛММ) in vitro (Moos et al., 1975; Starr & Offer, 1978; Safer & Pepe, 1980; Фрейдина и др., 1980; Подлубная, 1981; Podlubnaya et al., 1990). Показана способность С-белка связываться с актином (Moos et al., 1976; Moos et al., 1978; Moos, 1981; Фрейдина и др., 1980) и тайтином (Koretz et al., 1993; Soteriou et al., 1993; Fürst et al., 1992; Freiburg & Gautel, 1996). При снижении ионной силы ниже физиологических значений (μ ≤ 0.1).
Х-белок содержится в грубых препаратах С-белка, выделенного из миозина хроматографией на ДЕАЕ-сефадексе (Starr & Offer 1982). Был разработан метод разделения Х- и С-белков с помощью колоночной хроматографии на гидроксиапатите (Starr & Offer 1982; Starr et al., 1979). Молекула Х-белка представляет собой гибкую ниточку длиной около 30–35 нм и диаметром 3–4 нм (Bennett et al., 1985). Молекулярный вес Х-белка скелетных мышц кролика по данным ДСН-гель-электрофореза составляют и 145–152 кДа (Yamamoto & Moos 1983; Starr & Offer 1983), а по данным анализа нуклеотидной последовательности генов, кодирующего этот белок, составляют 126.5 кДа (Weber et al., 1993).
Аминокислотный состав характеризуется высоким содержанием пролина, что соответствует низкому (4%) содержанию α-спирали.
Н-белок выделен из скелетных мышц кролика, точнее – из грубых препаратов С-белка, хроматографией на гидроксиапатите (Starr & Offer, 1982; 1983). Он составляет 0.3–0.4% от миофибриллярной массы. Молекула Н-белка содержит одну полипептидную цепь с молекулярным весом по данным ДСН-гель-электрофореза ~ 69–74 кДа (Starr & Offer, 1983), а по данным другого метода ~ 86 кДа (Vaughan et al., 1993). Молекулярный вес Н-белка скелетных мышц цыпленка и человека по данным анализа нуклеотидной последовательности генов, кодирующего этот белок, составляет 58.4 кДа и 51.9 кДа, соответственно (Vaughan et al., 1993). Аминокислотный состав Н-белка сходен с аминокислотным составом С-белка, но отличается от последнего более высоким содержанием пролина и аланина и более низким содержанием аспарагиновой кислоты, валина и лизина (Yamamoto, 1984). Молекула Н-белка содержит около 4% α-спирали (Starr & Offer, 1983). Н-белок взаимодействует с миозином и с актином (Starr & Offer, 1983).
Белки семейства тайтина могут участвовать в сборке толстых нитей и стабилизации их структуры. Известно, что С-белок, также как и тайтин (Wang & Wright, 1988), появляется в мышечной клетке на ранних стадиях миофибриллогенеза (Bahler et al., 1985). Имеются также данные о влиянии тайтина, С-белка и Х-белка на формирование миозиновых нитей in vitro (Koretz, 1979; Miyahara & Noda, 1980; Koretz et al., 1982; Вихлянцев 2005). Вероятно, что С-белок (вместе с тайтином) участвует в сборке и стабилизации структуры толстых нитей. Исследования in vitro показали, что Н-белок влияет на связывание С-белка с миозином (Yamamoto, 1984). Показано связывание С-белка и тайтина с актином в условиях in vitro. Предполагается, что физиологическое значение этого связывания в саркомерах скелетных мышц заключается в регуляции актин-миозинового взаимодействия. В экспериментах in vitro показано, что тайтин и С-белок активируют АТФазу актомиозина. Это влияние может быть обусловлено как модифицирующим действием С-белка и тайтина на миозиновый мостик, так и прямым их взаимодействием с актином (Moos et al., 1978, 1980; Moos, 1981; Muhle-Goll et all., 2001; Moolman-Smook et al., 2002; Squire et al., 2003). Х-белок и Н-белок также могут быть вовлечены в процесс мышечного сокращения, ингибируя АТФ-азную активность актомиозина (Yamamoto, 1984, Вихлянцев, 2005). В поддержку предположения о регуляторной роли С-белка в сократительной активности являются данные о фосфорилировании С-белка сердечной мышцы. В ранних исследованиях было показано, что уровень фосфорилирования С-белка в интактном сердце коррелирует со скоростью релаксации мышцы (Hartzell & Glass, 1984). In vitro фосфорилирование сердечного С-белка приводило к снижению его активирующего эффекта на АТФазу АМ (Hartzell, 1985). На основе полученных данных автором было сделано предположение, что С-белок участвует в механизме расслабления сердечной мышцы. Однако результаты дальнейших исследований показали, что фосфорилирование С-белка модулирует процесс сокращения сердечной мышцы, увеличивая доступность миозиновых головок к актину (Weisberg & Winegrad, 1996, 1998; Kunst et al., 2000; McClellan et al., 2001; Kulikovskaya et al., 2003; McClellan et al., 2004; Flashman et al., 2004). Действительно, согласно последним данным фосфорилирование сердечного С-белка приводит к усилению сократительной активности мышцы (Stelzer et al., 2006).
Похожие работы
... эффект ?-сон индуцирующего пептида при гипокинетическом стрессе // Укр.биохим.-1991.-63.-№1.-С.34-37. 118.Механизмы развития стресса // Сб.статей.- Кишинев: Штиинца.- 1987.-222с. 119.Митюшина Н.В. Влияние энкефалинов на активность ферментов обмена регуляторных пептидов в головном мозге и периферических тканях крыс // дис. на соиск. степени.канд.биол.наук.- Пенза.-1999 120.Наркевич В.Б. ...
... крови в мокроте больного в период стихания процесса не является противопоказанием к назначению массажа по предлагаемой методике. Продолжая поиски возможностей более эффективного применения массажа при этой патологии, О.Ф-.Кузнецов (1979, 1980) предложил для больных хронической пневмонией, бронхиальной астмой и хроническим бронхитом новую методику и обосновал ее большую эффективность при равнении ...
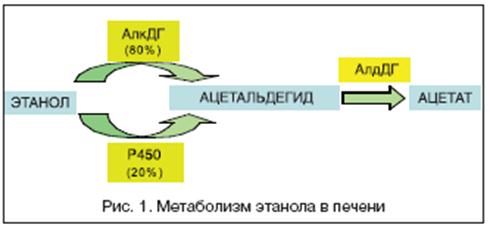


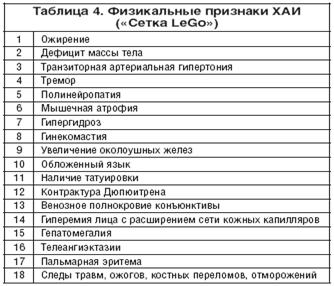
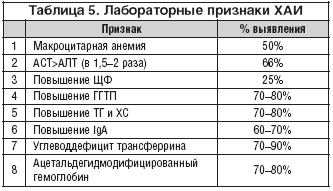
... , асцита, печеночной энцефалопатии). 5. Методом выбора при фульминантной форме ОАГ, а также у некоторых больных с алкогольным циррозом печени может быть трансплантация печени при условии минимум 6-месячной абстиненции [13]. 8. Клиническая фармакология гепатопротекторов В целом, ассортимент лекарственных средств, применяемых в комплексной терапии заболеваний печени и желчевыводящих путей, ...
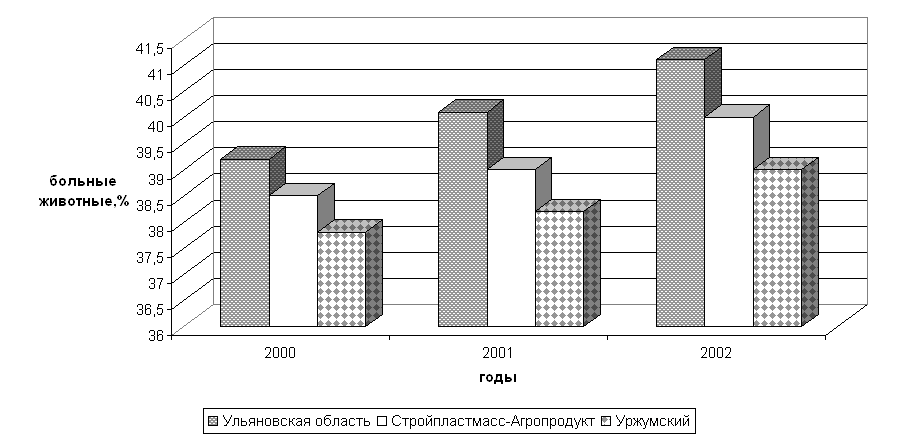
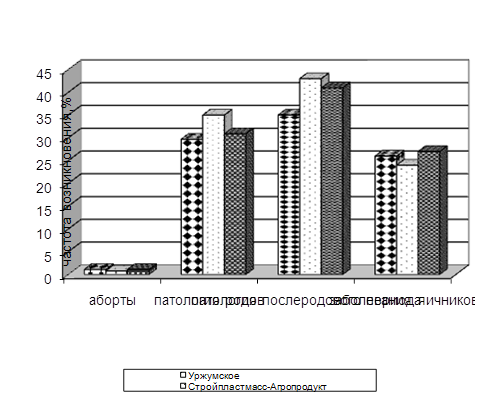
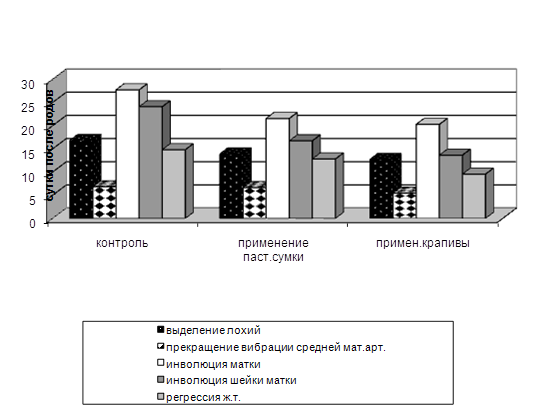
... изучить влияние вышеуказанных препаратов на морфологические, иммуно-биохимические и гемастазиологические параметры крови и течение послеродового периода высокопродуктивных молочных коров. 2.2.4 Профилактическая эффективность фитопрепаратов при патологии послеродового периода у высокопродуктивных молочных коров Патология родов и послеродового периода у коров широко распространена в молочном ...
0 комментариев