Системы транспорта ионов калия в митохондриях
Фармакологическая модуляция митохондриального
Структура митохондриального АТФ-зависимого
КДА и концентрирование белка методом обратного диализа с помощью полиэтиленгликоля (ПЭГ)
КДа, диализовали против 5 мМ Tris-HCl, pH 7.4
Концентрирование белка методом обратного диализа с помощью полиэтиленгликоля
Определение устойчивости к гипоксии крыс и адаптация к ней низкоустойчивых крыс
Изучение параметров АТФ-зависимого транспорта
Навигация
Системы транспорта ионов калия в митохондриях
Участие митохондриального АТФ-ингибируемого калиевого канала в адаптации животного к гипоксическому состоянию
61701
знак
8
таблиц
3
изображения
1.1. Системы транспорта ионов калия в митохондриях
Известно, что во внутренней мембране митохондрий локализуются ферменты синтеза АТФ, системы транспорта ионов и субстратов дыхания, а также переносчики электронов дыхательной цепи. При работе последних на мембране митохондрий создается потенциал (DY) около 190 мВ, который используется для синтеза АТФ. Существование высокого отрицательного DY создает предпосылки для входа в митохондрии катионов, среди которых основным является калий. Вход катионов К+ ведет к набуханию митохондрий и нарушению их структуры. Поэтому было предположено, что для поддержания объема митохондрий в них, наряду с системой потенциалозависимого входа, должна существовать система выхода катиона [44]. В дальнейшем это было подтверждено экспериментально [24, 56].
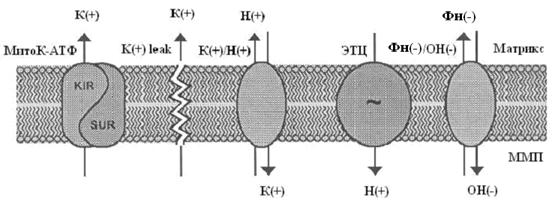
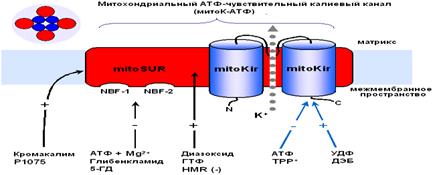
Так как скорость входа калия в митохондрии мала [14], первоначально полагали, что он осуществляется лишь благодаря диффузии иона через мембрану, проявляющейся в незначительных токах утечки [25]. Действительно, кажется маловероятным, что на внутренней мембране митохондрий, при наличии столь высокой электродвижущей силы, природа создала специфические каналы для калия. Однако, еще в 1981 году в лаборатории митохондриального транспорта Г. Д. Мироновой [4], а в дальнейшем и другими исследователями [17], из внутренней мембраны митохондрий был выделен селективный для калия канал, выполняющий функцию унипортера, причем антитела к этому белку-каналу специфически ингибировали работу данного канала, не влияя на другие функции митохондрий [6]. В настоящее время уже общепринято, что в митохондриях существует две системы транспорта калия: система унипортера, осуществляющего вход калия по электрохимическому потенциалу, и K+/H+-обменник, транспортирующий калий из митохондрий в обмен на Н+ [26,27]. То есть, в митохондриях существует так называемый калиевый цикл (рис 1).
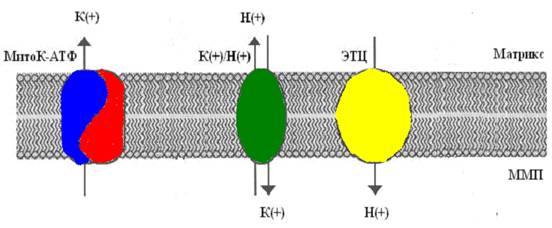
Рис 1. Митохондриальный калиевый цикл
(по Garlid and Paucek, 2003).
ЭТЦ – электрон-транспортная цепь; ММП – межмембранное пространство.
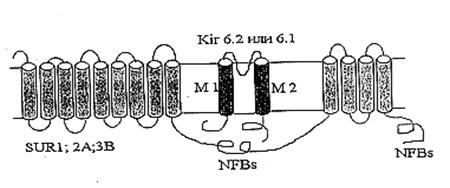
Первым компонентом этого цикла, выполняющим функцию унипортера, является белок-канал с молекулярной массой 55 кДа [3, 4, 43]. Этот белок, при встраивании в бислойные липидные мембраны (БЛМ), формирует селективные для ионов калия каналы. Позднее было обнаружено, что данный канал ингибируется АТФ [53]. Существование природного ингибитора канала вполне логично, так как закрытие канала предотвращает неконтролируемое набухание митохондрий. К настоящему времени митоКАТФ был обнаружен во внутренней мембране интактных митохондрий методом петч-кламп в нескольких лабораториях [12, 15, 20, 31, 45, 64].
Другим компонентом калиевого цикла является К+/Н+ - антипортер, который в настоящее время также выделен из внутренней мембраны митохондрий и молекулярная масса которого, по данным профессора Гарлида, равна 82 кДа [37]. Предполагается, что антипортер ответственен за поддержание объема митохондрий при увеличении скорости входа калия через внутреннюю митохондриальную мембрану. Открытие митоКАТФ кратковременно сдвигает баланс между К+ - унипортером и К+/Н+ - антипортером до тех пор пока скорость выхода К+ через последний не достигает скорости входа К+.
В настоящее время установлено, что митоКАТФ участвует не только в поддержании объема митохондрий, но и в адаптации животных к экстремальным воздействиям. По результатам, полученным на моделях гибернирующего суслика и адаптированных к холоду животных, в лаборатории митохондриального транспорта было высказано предположение об участии митоКАТФ в несократительном термогенезе [2,18]. Продолжение исследований в этом направлении может иметь значение для решения проблем анабиоза и гипотермического наркоза. Кроме того, в настоящее время стала широко изучаться роль митоКАТФ в кардиопротекции, что будет обсуждаться ниже.
1.2. Свойства и регуляция митохондриального АТФ-зависимого калиевого канала
Изучение свойств митоКАТФ показало, что он относится к семейству АТФ-зависимых калиевых каналов, впервые обнаруженных в цитоплазматических мембранах [46]. Основное общее свойство всех каналов, относящихся к этому семейству, состоит в том, что они ингибируются физиологическими концентрациями АТФ [31, 49, 53] и, являясь селективными для калия, обладают выпрямляющими свойствами, т.е. имеют разные скорости транспорта калия в зависимости от направления переноса ионов [31, 49].
Функционирование как митоКАТФ, так и цитоКАТФ зависит от редокс состояния активных групп белков-каналов. Установлено, что окислительно- восстановительные агенты модулируют работу митоКАТФ. Донор электронов п-диметиламиноэтилбензоат (ДЕБ) активирует митоКАТФ, а акцептор электронов - пеларгонидин ингибирует канал [29]. Эти реагенты влияют, вероятно, на SH-группы канала.
При гипоксии, как известно, в клетках значительно изменяется редокс баланс: наблюдается увеличение концентрации активных форм кислорода, изменение соотношений GSH/GSSG и НАД+/НАДН. Такие сдвиги могут привести к модификации тиоловых групп цистеинов.
Хорошо известно, что АФК образуются и накапливаются в организме при различных патологических состояниях, в частности, при ишемии. Однако, они могут образовываться в клетке при нормальном функционировании [48, 52] как побочные продукты аэробного метаболизма и играть значительную роль в работе дыхательной цепи [33], а также в процессах внутриклеточной сигнализации [22, 50].
В экспериментах со встроенным в БЛМ митоКАТФ было показано, что добавление в среду инкубации системы генерации радикала супероксид аниона – ксантин/ксантин оксидазы, приводило к активации канала уже в течение первой минуты инкубации [64]. При этом вероятность открытого состояния митоКАТФ возросла приблизительно в три раза. По-видимому, действие свободных кислородных радикалов на канал опосредовано различными механизмами и направлено на его сульфгидрильные группы.
Другим универсальным фактором регуляции метаболизма является оксид азота II (NО).
Недавно стало известно, что синтез NO может происходить локально в митохондриях, возможно, за счет существования специфичной митохондриальной NO-синтетазы – mtNOS [59]. Было показано, что образованный в тканях NO обладает активирующим действием на митоКАТФ [15, 36, 57]. Так как дыхательная цепь митохондрий является постоянным источником супероксид аниона, который легко вступает в реакцию с NO, вполне вероятно, что в митохондриях происходит образование пероксинитрита ONOO- [38]. Предполагают, что активирующий эффект NO и пероксинитрита (ONOO-)на митоКАТФ осуществляется через активацию протеинкиназы С [35].
Похожие работы
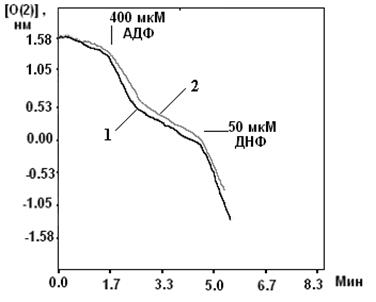
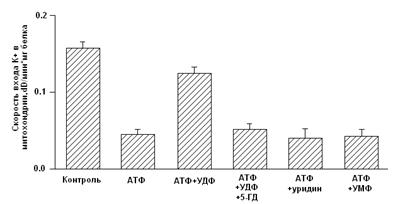
... м, в течение 10-15 мин. Группа низкоустойчивых (НУ) животных выдерживала эту высоту только в течение 1-1.5 мин. Глава 5. Результаты и обсуждения 5.1 Параметры функционирования митоКАТФ канала у крыс с различной резистентностью, а также у животных, адаптированных к гипоксии В этом разделе работы исследовались такие показатели, как дыхание МХ, скорость АТФ-зависимого К+ транспорта, ...
0 комментариев