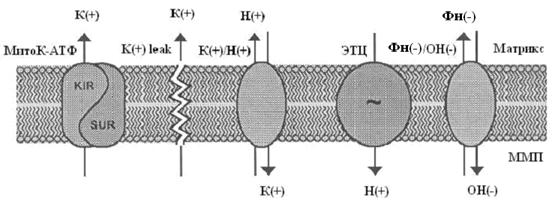
Системы транспорта ионов калия в митохондриях
Фармакологическая модуляция митохондриального
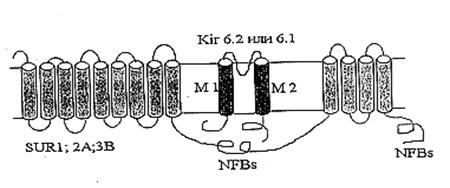
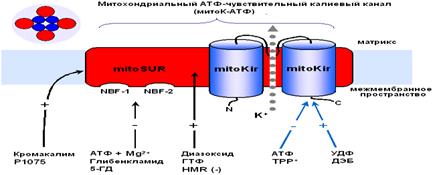
Структура митохондриального АТФ-зависимого
КДА и концентрирование белка методом обратного диализа с помощью полиэтиленгликоля (ПЭГ)
КДа, диализовали против 5 мМ Tris-HCl, pH 7.4
Концентрирование белка методом обратного диализа с помощью полиэтиленгликоля
Определение устойчивости к гипоксии крыс и адаптация к ней низкоустойчивых крыс
Изучение параметров АТФ-зависимого транспорта
Навигация
Фармакологическая модуляция митохондриального
Участие митохондриального АТФ-ингибируемого калиевого канала в адаптации животного к гипоксическому состоянию
61701
знак
8
таблиц
3
изображения
1.3. Фармакологическая модуляция митохондриального
АТФ-зависимого калиевого канала
Классификация митоКАТФ как селективного К+ канала, напоминающего изоформу цитоКАТФ, следует, в основном, из того, что оба канала имеют общие активаторы и ингибиторы. В настоящее время обнаружено большое количество подобного рода фармакологических агентов [51]. Эти данные обобщены в таблице 1.
Таблица 1. Модуляторы цитоплазматического и митохондриального КАТФ каналов.
| Модуляторы | митоКАТФ | митоКАТФ и цитоКАТФ | цитоКАТФ |
| Активаторы | диазоксид никорандил BMS-180448 BMS-191095 ДЕБ Тестостерон УДФ | кромакалим пинацидил Р-1060 силденафил изофуран EMD60480 априкалим P-1075 β- эстрадиол | МСС-134 |
| Ингибиторы | 5-HD МСС-134 | глибенкламид | HMR1098(1833) Глимепирид |
Селективность действия модулятора зависит от типа изучаемых клеток, условий эксперимента и, в основном, от используемых концентраций. Диазоксид и никорандил, например, активируют не только митоКАТФ, но в более высоких концентрациях также активируют и цитоКАТФ [16,58]; 5-HD помимо митоКАТФ ингибирует при низких значениях рН цитоКАТФ, активируемый АДФ [47].
Известно, что MCC-134, аналог априкалима, в поджелудочной железе ингибирует, а в гладкой мускулатуре активирует КАТФ каналы. Недавно было показано, что этот препарат ингибирует митоКАТФ, но активирует цитоКАТФ в кардиомиоцитах.
Недавно, в лаборатории митохондриального транспорта было показано, что дифосфонуклеотиды являются активаторами канала, причем наиболее выраженный эффект наблюдался при активации канала УДФ.
Было показано, что дифосфонуклеотиды, такие как АДФ и ГДФ активируют реконструированную в БЛМ канальную субъединицу [29,43]. Данные по активирующему действию ГДФ в дальнейшем были подтверждены и другими исследователями [12]. Недавно было показано, что в ряду дифосфонуклеотидов наиболее эффективным является уридиндифосфат (УДФ) [42]. Этот фосфонуклеотид в микромолярных концентрациях активирует митоКАТФ. Для активации цитоКАТФ нужны значительно большие концентрации УДФ [8]. Реконструированный в БЛМ канал (митоKIR), после ингибирования АТФ, полностью реактивируется 20 мкМ УДФ. В интактных митохондриях, где присутствуют обе субъединицы, канал реактивируется теми же концентрациями УДФ, и эта активация снимается глибенкламидом и 5-НD [42]. Следовательно, участок связывания УДФ локализуется на канальной субъединице.
Функцию метаболических регуляторов могут выполнять также гормоны. Еще на первых этапах исследования митоКАТФ в лаборатории митохондриального транспорта обнаружено, что прогестерон ингибирует его канальную субъединицу. В настоящее время появились данные о том, что женский половой гормон b-эстрадиол является, вероятно, активатором митоКАТФ, так как он обладает кардиопротекторным действием, которое снимается 5-НD [61].
Мужской половой гормон – тестостерон, также оказывал как активирующее митоКАТФ, так и кардиопротекторное действие [20]. При измерении активности митоКАТФ по флуоресценции флавопротеидов в митохондриях сердца, а также методом петч-кламп по регистрации электрических характеристик мембран митопластов было установлено, что тестостерон вызывал окисление флавопротеидов и активацию каналов. Оба эффекта зависели от присутствия в среде К+ и были чувствительны к АТФ. Показано, что тестостерон оказывал эффект только на митоКАТФ и не влиял на цитоКАТФ [20].
Каналы, образованные в БЛМ белком с молекулярной массой 55 кДа, регулируются АТФ, как было ранее показано [3], который способен значительно снижать проводимость модифицированной белком мембраны. Ионы Mg2+ не оказывали значительного влияния на ингибирование канала АТФ.
В интактных митохондриях можно также наблюдать АТФ-зависимый транспорт калия, используя модель энергозависимого входа калия. АТФ в присутствии MgCl2 способен был подавлять энергозависимый вход калия в митохондрии. Однако, в отличие от данных, полученных при реконструкции этого белка в искусственные мембраны, действующая концентрация АТФ значительно ниже. Другим отличием является абсолютная необходимость Mg2+ для ингибирования АТФ в случае интактных митохондрий, в то время как при реконструкции белка с Мr55 кДа в БЛМ, Mg2+ не оказывает существенного влияния на эффект ингибирования калиевого канала АТФ.
Различия в действии Mg2+, возможно, объясняются тем, что Mg2+ - связывающий участок локализован на регуляторной субъединице канала. Связывание регуляторной субъединицы с магнием увеличивает, вероятно, сродство к АТФ каналообразующей субъединицы.
Ингибирование нуклеотидом, вероятнее всего, осуществляется за счет его связывания в специфическом участке, а не в результате фосфорилирования белка, хотя таковое может иметь место. Интересно, что присутствие магния не является необходимым условием для ингибирующего эффекта АТФ при реконструкции митоKir (белка с Мr 55 кДа) в искусственные мембраны, следовательно, Mg2+ проявляет свой эффект только в интактных митохондриях, то есть при наличии двух субъединиц.
Так как ингибирующий эффект АТФ проявляется и при реконструкции только каналоформирующей субъединицы (митоKir) в БЛМ и липосомы, логично предположить, что нуклеотидсвязывающий участок расположен на каналообразующей субъединице, однако регуляторная субъединица (митоSUR) повышает сродство каналов к нуклеотиду.
В исследовании по регуляции транспорта калия адениновыми нуклеотидами с использованием модели выхода ионов K+ из митохондрий в присутствии разобщителя окислительного фосфорилирования было обнаружено, что ингибитор адениннуклеотидтранслоказы ― атрактилозид в концентрации 1 мкМ полностью предотвращал ингибирующее действие АТФ. Следовательно, можно предположить, что для ингибирования канала АТФ должен транспортироваться внутрь митохондрий, где и происходит его связывание с каналом. Исходя из этого, представляется, что нуклеотидсвязывающий участок митохондриального АТФ-чувствительного калиевого канала располагается на обращенной в сторону матрикса части белка-канала.
Похожие работы
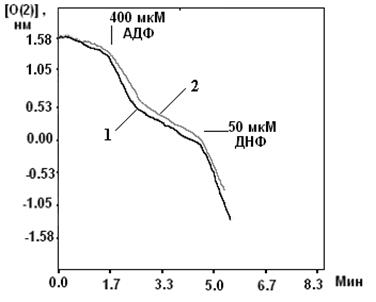
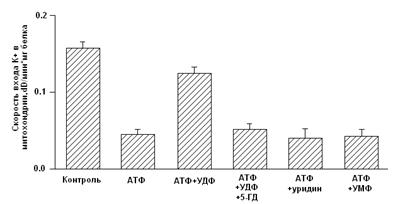
... м, в течение 10-15 мин. Группа низкоустойчивых (НУ) животных выдерживала эту высоту только в течение 1-1.5 мин. Глава 5. Результаты и обсуждения 5.1 Параметры функционирования митоКАТФ канала у крыс с различной резистентностью, а также у животных, адаптированных к гипоксии В этом разделе работы исследовались такие показатели, как дыхание МХ, скорость АТФ-зависимого К+ транспорта, ...
0 комментариев