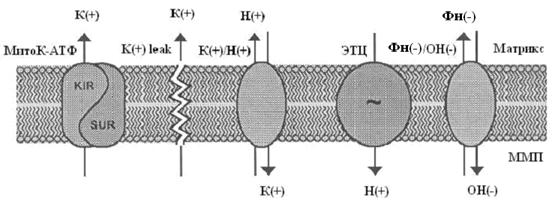
Системы транспорта ионов калия в митохондриях
Фармакологическая модуляция митохондриального
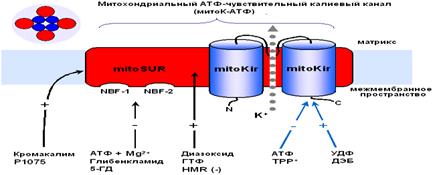
Структура митохондриального АТФ-зависимого
КДА и концентрирование белка методом обратного диализа с помощью полиэтиленгликоля (ПЭГ)
КДа, диализовали против 5 мМ Tris-HCl, pH 7.4
Концентрирование белка методом обратного диализа с помощью полиэтиленгликоля
Определение устойчивости к гипоксии крыс и адаптация к ней низкоустойчивых крыс
Изучение параметров АТФ-зависимого транспорта
Навигация
Структура митохондриального АТФ-зависимого
Участие митохондриального АТФ-ингибируемого калиевого канала в адаптации животного к гипоксическому состоянию
61701
знак
8
таблиц
3
изображения
1.4. Структура митохондриального АТФ-зависимого
калиевого канала
Структура цитоКАТФ и ген ее кодирующий к настоящему времени известны. Установлено, что этот канал состоит из двух субъединиц: канальной и регуляторной. Канальная субъединица имеет Мr 43-46 кДа и формирует АТФ-зависимый калиевый канал, который не регулируется специфическими модуляторами канала. Регуляторная субъединица имеет Мr от 60 до 174 кДа, в зависимости от ткани, и придает каналу чувствительность к модуляторам [7].
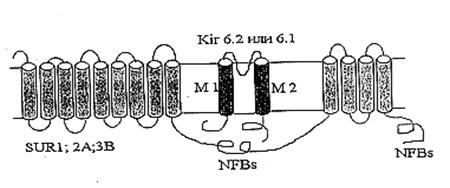
Несмотря на то, что свойства, регуляция и физиологическое значение митоКАТФ в настоящее время широко изучаются, солюбилизация его из митохондриальных мембран и изучение структуры проводятся лишь в отдельных лабораториях. Так как митоКАТФ имеет ряд общих свойств с цитоКАТФ и регулируется общими регуляторами, было предположено, что по структуре он близок к цитоплазматическому каналу [43] и состоит из канальной субъединицы [42] и регуляторной [10]. МитоКАТФ, также как и клеточный, является, по всей вероятности, гетеромультимером, состоящим из калиевого канала, белка с молекулярной массой 55 кДа, который имеет выпрямляющие свойства и который, по аналогии с цитоплазматическим каналом, назвали митоKIR [42] и рецептора, чувствительного к сульфонилмочевинам и поэтому названного митоSUR (Рис. 2) [10, 43].
Рис 2. Рабочая модель структуры митохондриального АТФ-зависимого калиевого канала (по Mironova et al., 2004)
Белок, выделенный из внутренней мембраны митохондрий печени крысы, который формирует в БЛМ АТФ-зависимые каналы и имеет Мr 55 кДа (митоKIR), не ингибируется глибенкламидом и специфическим ингибитором митоКАТФ 5-HD, и не активируется кромакалимом и диазоксидом. В то же время эти препараты влияли на АТФ-зависимый калиевый транспорт в изолированных митохондриях [42].
Константа ингибирования АТФ на реконструированном в БЛМ митоKIR была значительно выше, чем в интактных митохондриях и Mg2+ для этого ингибирования не требовался. В то же время, для ингибирования фосфонуклеотидом канала, локализованного в интактных митохондриях, присутствие ионов магния было необходимо [42]. Те же различия были обнаружены при сравнительном изучении влияния АТФ и сульфонилмочевины на цитоKIR и целый цитоКАТФ [62]. Целый цитоКАТФ ингибировался значительно меньшими концентрациями АТФ, чем канальная субъединица и был чувствителен к сульфонилмочевине и активаторам, в то время как канальная субъединица такой чувствительности не проявляла. Из этих данных был сделан вывод, что основной участок связывания АТФ в цитоКАТФ локализован на канальной субъединице, а регуляторная субъединица повышает сродство канальной субъединицы к АТФ и обеспечивает чувствительность целого канала к активаторам и ингибиторам [62].
Приведенные выше результаты исследований митоКАТФ подтверждают предположение о том, что белок с Мr 55 кДа является канальной субъединицей целого митоКАТФ. Однако, для более прямого подтверждения этого предположения нами были получены антитела (АТ) на этот белок и показано, что они ингибируют АТФ-зависимый калиевый транспорт в интактных митохондриях, не влияя при этом на другие функции митохондрий, такие как окислительное фосфорилирование и транспорт кальция [6]. Функцию регуляторной субъединицы выполняет, вероятно, белок с Мr 63 кДа, связывающийся с меченым глибенкламидом [10].
1.5. Нарушения гомеостаза, вызванные гипоксией и механизмы его восстановления
Известно, что при гипоксии недостаток кислорода приводит к восстановлению переносчиков дыхательной цепи [1]. Восстановление переносчиков I и III комплексов цепи, приводит к увеличению образования АФК, которые активируют свободнорадикальные процессы в клетке [19, 32].
Можно выделить две основные физиологические роли свободнорадикального окисления в организме. Во-первых, этот процесс имеет место при катаболизме многих липидов и белков, что облегчает дальнейшее действие фосфолипаз и протеаз, сродство которых намного выше именно к окисленному субстрату [63], а также при синтезе физиологически активных веществ липидной природы (лейкотриены, тромбоксаны, простагландины). Во-вторых, АФК участвуют в начальных этапах клеточной сигнализации (редокс–сигнализация) в условиях стресса, гипоксии, воспаления, воздействия высоких и низких температур, физической нагрузки, и других патологических состояниях. Характер клеточного ответа будет зависеть от продолжительности и интенсивности воздействия вышеперечисленных факторов. При умеренном воздействии формируется неспецифический ответ, повышающий адаптацию организма к новым условиям. При воздействии высокой интенсивности, например, при глубокой гипоксии наступает некроз тканей, в том числе и за счет повреждающего действия АФК, активирующих перекисное окисление липидов и других биологических молекул [5].
Механизм протекторного действия заключен, по-видимому, в окислительно-восстановительных модификациях сульфгидрильных групп сенсорных белков, что приводит к активации тирозинкиназного пути клеточного ответа [5].
Одним из важнейших следствий инициации редокс-сигнализации и АФК-опосредованной передачи сигнала является активация ядерных факторов транскрипции, которые находятся в неактивном состоянии до тех пор, пока в их молекуле не произойдет отщепление ингибиторного домена. После этого, ядерные факторы транскрипции оказываются способными индуцировать многочисленные гены. Важную роль в настоящее время придают таким факторам транскрипции, как NF-kB [21], AP-1[41].
Среди известных к настоящему времени белков, которые синтезируются в ответ на редокс-сигнал от адаптирующего фактора, наибольшее значение имеет, прежде всего, ряд неспецифических молекул, таких как ферменты антиоксидантной защиты, белки семейства HSP и другие белки срочного ответа, которые могут синтезироваться в ответ на гипоксию, стресс, ишемию, реперфузию и т.д. [41]. Кроме того, синтезируются специфические молекулы с шапероновой активностью по отношению к конкретным белкам клетки, как показано, например для специфического шаперона Са-насоса саркоплазматического ретикулума [13], либо белки, специфически синтезирующиеся в ответ на действие конкретного фактора, например, в ответ на гипоксию [28].
Следует отметить, что на сегодняшний день одним из наиболее эффективных способов предотвращения повреждений тканей, вызванных гипоксическим состоянием, является интервальная гипоксическая тренировка. До настоящего времени считалось, что основной механизм адаптационного эффекта всех видов гипоксической тренировки обусловлен активацией антиоксидантных ферментов, усиливающих защиту организма от воздействия активных форм кислорода [28,41]. Недавно в лаборатории Скулачева было показано, что незначительное снижение мембранного потенциала (~ на 13%) ведет к существенному (до 80%) уменьшению продукции супероксид-анионов [34]. Следует отметить, что в митохондриях образуется до 95% супероксид-анионов клетки [11,54,60].
Ранее было высказано предположение, что активация митоКАТФ и увеличение экспрессии гена, кодирующего этот белок, играют роль в формировании устойчивости организма к кислородному голоданию [39,55,65], однако прямых доказательств этому предположению получено не было, поэтому в данной работе была поставлена цель – выяснить роль митоКАТФ в адаптации к гипоксии.
Глава 2. Материалы и методы
2.1. Выделение белка с молекулярной массой 55 кДа из митохондрий печени крыс
В работе использовали самцов крыс линии Вистар.
Для выделения белка проводили выделение митопластов, экстракцию белков из митопластов, отделение водорастворимых белков от нерастворимых в воде. Дальнейшее разделение водорастворимых белков осуществляли методом ионообменной хроматографии, с повторным проведением хроматографии фракций, где был обнаружен белок с Мr 55 кДа. Фракции после хроматографии проверяли с помощью SDS-электрофореза и, если было необходимо, на приборе для определения проводимости бислойных липидных мембран. Далее проводили нативный форез фракций после повторной хроматографии, в которых с помощью SDS-фореза был обнаружен белок с Мr
Похожие работы
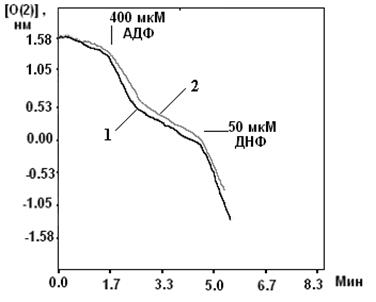
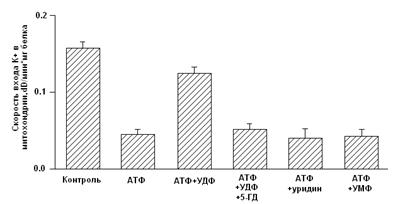
... м, в течение 10-15 мин. Группа низкоустойчивых (НУ) животных выдерживала эту высоту только в течение 1-1.5 мин. Глава 5. Результаты и обсуждения 5.1 Параметры функционирования митоКАТФ канала у крыс с различной резистентностью, а также у животных, адаптированных к гипоксии В этом разделе работы исследовались такие показатели, как дыхание МХ, скорость АТФ-зависимого К+ транспорта, ...
0 комментариев