РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
РОЛЬ БЕЛКОВ В ДИНАМИЧНОСТИ ЛИПИДНОГО БИСЛОЯ
МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
ЛИПИДЫ ВНЕШНЕЙ ЗОНЫ МЕМБРАН МОЗГА
Локализация ганглиозидов в головном мозге
Участие ганглиозидов в дифференциации клеток
Межклеточное гликозирование ганглиозидов
О-ацетилирование ганглиозидов – один из возможных механизмов изменения их структуры
ИЗМЕНЕНИЕ СОСТАВА ЛИПИДОВ В ОНТОГЕНЕЗЕ
Навигация
РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
Липиды центральной нервной системы и структура клеточных мембран
82146
знаков
8
таблиц
16
изображений
1. РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
В мембранах головного мозга имеет место цикл деацилирование – реацилирование, при котором происходит замена жирных кислот в молекуле фосфолипидов, в то время как другие компоненты молекулы остаются неизменными. Этот ацилобменный механизм является особенно важным для включения тех или иных жирных кислот во второе положение остатка глицерина, и его рассматривают как средство локального регулирования физических и функциональных свойств мембран. Существенную роль играет и переход диацильных форм фосфолипидов в моноацильные и обратно. Все это оказывает влияние на такие мембранные процессы, как проницаемость для различных веществ, транспорт ионов и т.д.
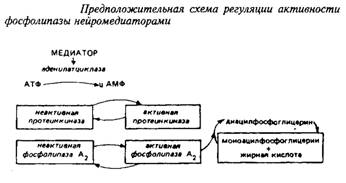
Ацилобменные реакции имеют прямое отношение ко многим процессам, влияя на активность ряда ферментов, на синтез простагландинов и чувствительность фоторецепторов. Некоторые исследователи связывают ферментативное деацилирование – реацилирование с эффектом синаптической передачи. Так, под влиянием норадреналина в синаптосомах происходит активирование фосфолипазы А2, отщепляющей жирную кислоту во втором положении глицерофосфолипида. Таким образом, нейромедиатор модифицирует обмен фосфолигавдов в синаптических мембранах путем вовлечения в этот процесс реакций деацилирования. Предложена следующая схема регуляции активности ацилобменного цикла нейромедиаторами.
2. ОРГАНИЗАЦИЯ ЛИПИДОВ В МЕМБРАНЕ
Образование липидных молекул в ходе эволюции и выбор именно этих молекул в качестве строительных блоков мембран сыграли решающую роль в возникновении жизни. Липидам принадлежит жизненно важная роль в клетке. Следующие особые физико-химические свойства липидов определяют их роль в построении мембран:
1. Сочетание гидрофильных и липофильных свойств в структуре одной молекулы, их амфифильность.
2. Способность липидов четко ориентироваться на границе раздела фаз, так что полярные группы направлены в водные среды, а неполярные экранированы от них.
3. Способность липидов самопроизвольно упаковываться в прочные, плотные мономолекулярные слои или пленки, устойчивые к сжатию. Плотность такой упаковки зависит от рН, температуры и молекулярной организации липидов. Такие плотные слои создают определенный барьер для диффузии молекул.
4. Способность липидов агрегировать в хорошо упорядоченные сферические, цилиндрические, ламеллярные мицеллы. В мицеллах липиды ориентированы таким образом, чтобы максимальное число полярных групп находилось в контакте с водой, а гидрофобная часть была максимально удалена от контакта с ней.
Способность липидов образовывать прочные мономолекулярные слои лежит в основе молекулярной организации мембран. Более 60 лет назад было высказано предположение, что в основе мембран лежит бимолекулярный слой липидов.
В бимолекулярном липидном слое гидрофобные цепочки молекул липидов направлены друг к другу и внутренность бислоя совершенно гидрофобна, а гидрофильные части образуют поверхности внутреннего и внешнего монослоев, открытые для разнообразного рода взаимодействий.
Липидный состав мембран нервной ткани и распределение липидов по слоям генетически детерминированы. Наружный и внутренний монослои липидов характеризуются планарной и поперечной микрогетерогенностьюу что создает асимметричность мембран. Существует несколько механизмов, поддерживающих асимметричное распределение липидов в мембране. Один из них связан с термодинамической вероятностью размещения липид-ных молекул с учетом их стереоконфигурации, заряда и гидратации полярных групп. Так, основная часть фосфатидилхолина, сфингомиелина, полифосфоинозитидов, холестерина, церебро-зидов и сульфатидов локализована в наружном слое, а амино-фосфолипиды находятся во внутреннем, цитоплазматическом слое. Неодинакова степень ненасыщенности монослоев: во внутреннем обнаруживается 2/3 двойных связей в жирных кислотах липидов, а в наружном – только 1/3.
Асимметрия бислоя является фактором, обеспечивающим создание градиента кривизны, складок, сморщиваний, отшнуровки части мембраны в виде везикул что существенно для обеспечения межклеточных взаимодействий.
Другой механизм поддержания асимметрии бислоя реализуется за счет различий ионного состава вне- и внутриклеточной среды, что вносит вклад в создание и поддержание изгибов мембраны.
Асимметрия бислоя обеспечивается также ферментами ли-пидного обмена, к ним прежде всего относятся липазы, ферменты обмена холестерина и метилазы фосфатидилэтанолами-на. Метилирование фосфатидилэтаноламина с превращением его в фосфатидилхолин осуществляется в два этапа и происходит в разных слоях липидкого матрикса. Образование мономе-тилфосфатидилэтаноламина под влиянием метилтрансферазы I осуществляется во внутреннем слое, где и локализован фермент. Монометил фосфатидилэтаноламин переходит из цитоплаз-матического слоя на внешний, где под действием метилтрансферазы II завершается его превращение в фосфатидилхолин. Фактически осуществляется так называемый ферментативный флип-флоп.
Этот транслокационный процесс меняет жидкостность мембраны и рассматривается как фактор, стимулирующий функционально важные процессы в мембране: связывание рецепторов с лигандами, Са* – вызванное освобождение медиаторов из си-наптических окончаний, активирование ЛТФаз.
Асимметричность билипидного слоя может поддерживаться транспортом липидов: спонтанным, везикулярным или с участием липидпереносящих белков. Липидпереносящие белки различной степени специфичности «стоят на страже» асимметрии мембран, перенося липиды только в наружный или только во внутренний слой. Перенос липидных молекул осуществляется в виде комплексов с белками-переносчиками.
Похожие работы
... в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой. Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение ...
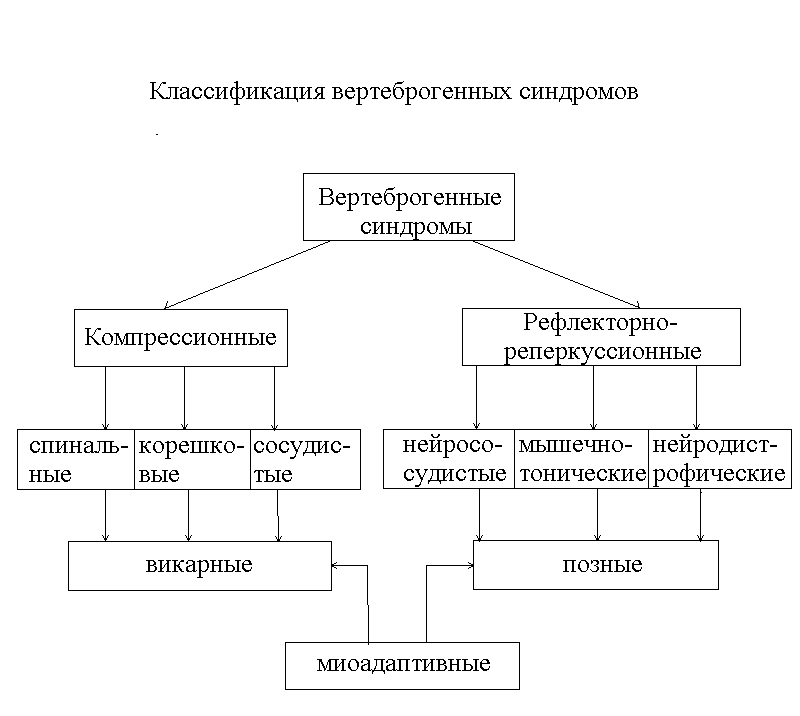
... , лептоспироз и др.) и вторичными (вертеброгенные, после детских экзантемных инфекций, инфекционного мононуклеоза, при узелковом периартериите, ревматизме и др.). По патогенезу и патоморфологии заболевания периферической нервной системы подразделяются на невриты (радикулиты), невропатии (радикулопатии) и невралгии. Невриты (радикулиты) – воспаление периферических нервов и корешков. По характеру ...

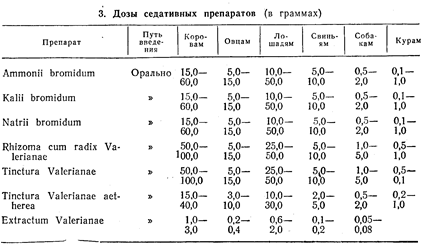

... , обволакивающие, адсорбирующие и др.). В этих же направлениях влияют местноанестезирующие вещества, выключающие отдельные звенья чувствительных нервов. Подавить боль можно, применяя вещества, угнетающие центральную нервную систему. Все эти средства используют при соответствующих показаниях. Но при болевых синдромах, являющихся следствием воспалительных процессов в организме, исключительно ...
... составляют тела клеток, дендриты и немиелинизированные аксоны, организованные в комплексы, которые включают бесчисленное множество синапсов и служат центрами обработки информации, обеспечивая многие функции нервной системы. Белое вещество состоит из миелинизированных и немиелинизированных аксонов, выполняющих роль проводников, передающих импульсы из одного центра в другой. В состав серого и белого ...
0 комментариев