РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
РОЛЬ БЕЛКОВ В ДИНАМИЧНОСТИ ЛИПИДНОГО БИСЛОЯ
МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
ЛИПИДЫ ВНЕШНЕЙ ЗОНЫ МЕМБРАН МОЗГА
Локализация ганглиозидов в головном мозге
Участие ганглиозидов в дифференциации клеток
Межклеточное гликозирование ганглиозидов
О-ацетилирование ганглиозидов – один из возможных механизмов изменения их структуры
ИЗМЕНЕНИЕ СОСТАВА ЛИПИДОВ В ОНТОГЕНЕЗЕ
Навигация
ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
Липиды центральной нервной системы и структура клеточных мембран
82146
знаков
8
таблиц
16
изображений
3. ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
Строгая организованность липидного слоя мембраны не лишает его большой динамичности, которая возникает из-за передвижения липидных молекул в пределах мембраны, т.е. за счет интрамолекулярных движений липидов в пределах бислоя. Известно по крайней мере четыре типа интрамолекулярных движений липидов в пределах мембраны: латеральная диффузия, вращательная диффузия, вертикальные колебания и упоминавшийся выше так называемый флип-флоп.
Для большинства липидов скорость латеральной диффузии ощутима. Коэффициент латеральной диффузии для липидов Ю – см/с, а для белков намного ниже – 10~ ш/с. Вращательная диффузия молекул осуществляется также легко. Скорость же флип-флопа очень низка. Особенно медленно флип-флоп происходит в чисто липидных везикулах. Даже в присутствии липидпереносящих белков перемещение из наружного слоя во внутренний занимает более 4 часов, а перемещение в обратную сторону – более 10 часов. Не ускоряет флип-флоп повышение температуры до 80°С Это движение фосфолипидов усиливается под влиянием окисленных липидов, лизолецити-на. Как правило, холестерин подвергается более быстрому флип-флопу, чем фосфолипиды. Следует отметить, что не только флип-флоп запускает функционально важные события в мембране. Латеральная и вращательная диффузия липидов оказывает регулирующее влияние на активность мембранных белков.
4. ФАЗОВЫЕ ПЕРЕХОДЫ ЛИПИДОВ В МЕМБРАНЕ
На все типы молекулярных движений липидных молекул сильное влияние оказывает структура, в которой в данный момент находится липвды бислоя – гелеобразная или жидко-кристаллическая.
Липиды обладают замечательным свойством – способностью к фазовым переходам в физиологических условиях. При определенных температурах, строго характерных для каждого вида липидов, липидные мицеллы могут быть в «твердом» кристаллическом, организованном, гелеобразном состоянии или в «жидком», мезофазном, так называемом жидко-кристаллическом состоянии. Жидкие кристаллы – это анизотропные жидкости, так как оптически они сходны с кристаллами, проявляя разные свойства в разных направлениях, а механически сходны с жидкостью, они текут в зависимости от вязкости.
От состояния липидов в мембране зависит уровень молекулярной организации. Липиды в кристаллическом состоянии могут быть упакованы в кубический или гексагональный кристалл. Жидко-кристаллическая организация липидов в мембране очень разнообразна – это так называемые нематики, смек-тики, холестерики.
Нематики – наименее упорядоченная организация жидко-кристаллического состояния липидов. Молекулы нематика при умеренной температуре стремятся ориентироваться вдоль одного направления. В нематике очень многие молекулы одинаково ориентированы, их продольные оси параллельны друг другу, но такие области существуют недолго и границы их размыты. Области с одинаковой ориентацией молекул непрерывно рождаются и исчезают, что зависит от многих условий – внешних границ, включений и различных воздействий. Магнитное и электрическое поля ориентируют молекулы нематика, причем выстраивают молекулярные оси параллельно своему направлению.
Смектики – похожи на мыльные пленки, они более организованы, чем нематики, их молекулы образуют слои. В каждом индивидуальном слое молекулы передвигаются вдоль плоскости, все плоскости слоев находятся на одном и том же расстоянии. Смектики очень пластичны. Так, смектик в нативной мембране при охлаждении превращается в нематик.
Спиральная упаковка молекул вносит новое в ориентацию оптической оси жидкого кристалла. У холестериков – слоистое строение с различным шагом спирали. Холестерическую спираль обозначают нередко как твист-ориентацию. Разбавление холестерика и увеличение шага спирали приводит к нема-тику. Оптическая активность холестериков очень велика, они избирательно отражают свет в зависимости от температуры, механической нагрузки, примесей, электромагнитных полей.
Жидкие кристаллы, сочетая в себе упорядоченность твердого тела и подвижность жидкости, отличаются высокой чувствительностью к внешним воздействиям, температуре, примесям, свету, внешним полям, они очень пластичны и очень долго хранят информацию. Эти свойства приобретают первостепенное значение в мембранах нервной ткани, где изменения электрических свойств лежат в основе проведения возбуждения.
Фазовый переход липидов является эндотермическим процессом, сопровождающимся изменением энтропии и энтальпии. Липидным структурам присущ лиотропный мезоморфизм и термотропный мезоморфизм. Оба свойства связаны между собой. Фазовый переход липидов «гель – жидкий кристалл» осуществляется при температуре, значение которой зависит от содержания воды в системе. Оно минимально, если общее содержание воды превышает то количество, которое могут связать липидные структуры. В то же время при температуре выше критической липиды могут находиться в упорядоченном состоянии при недостатке воды. Перекисное окисление липидов, увеличивающее содержание воды в бислое, существенно влияет на фазовое состояние мембраны.
Термотропные фазовые переходы липидов в мембране происходят в сравнительно широком температурном интервале. Это обусловлено тем, что в бислое одна фаза обязательно возникает в матриксе другой. Сосуществование в липидном бислое двух фаз устанавливает между ними сложное равновесие, приводя к снижению степени кооперативное™ перехода. Обычно кооперативные фазовые переходы липидов в мембране затрагивают несколько сотен молекул. В нативной мембране постоянно находится большое число кооперативных единиц той или иной фазы. Этот полиморфизм является мощным регулятором транспортных систем мембраны.
Следует отметить, что на температуру фазового перехода большое влияние оказывают структура липидной молекулы, длина углеводородного скелета, наличие цис- и транс-двойных связей, структура полярных групп.
При переходе в жидко-кристаллическое состояние имеет место несколько одновременных событий: возрастает подвижность полярных групп липидов, увеличивается вращательная подвижность жирнокислотных радикалов относительно С–С-связей, увеличивается скорость латеральной диффузии. Это приводит к изменению геометрических размеров бислоя из-за латерального расширения площади, занимаемой каждой молекулой липида. Например, площадь, занимаемая 2С]6-фосфати-дилхолином, меняется от 0,49 до 0,58 нм, среднее расстояние между цепями увеличивается от 0,49 до 0,52 нм, а толщина углеводородного скелета уменьшается почти на 0,5 нм, т.е. латеральное расширение компенсируется утончением слоя. Гидрофобный объем мембраны увеличивается примерно на 1,5%.
В результате этих и ряда других изменений состояния липидов в мембране создаются особые условия для проникновения гидрофобных вешеств, изменения работы ионных каналов, внедрения в мембрану различных белков.
Микрогетерогенность бислоя и образование в нем кластеров молекул липидов способствует проявлению такого явления, как разделение фаз в мембране. Латеральное разделение липидных молекул в плоскости бислоя – важная особенность мембраны. Особая сегрегирующая роль в мембране принадлежит холестерину. При низких концентрациях его в мембране происходит латеральное разделение фосфолипид-холе-стсриновых комплексов и свободных молекул фосфолипидов. При этом холестерин взаимодействует в первую очередь с теми молекулами фосфолипидов, которые имеют низкую температуру фазового перехода. Благодаря этому в бислое будут существовать области только жидкие и только твердые, а также области, где обе фазы сосуществуют. Наличие таких жидких и твердых областей в пределах мембраны изменяет ее сжимаемость, что сказывается на глубине погружения мембранных белков и на эффективности работы мембранных насосов.
Необходимо отметить, что кроме сегрегирующего холестерин проявляет и другое важное влияние на структуру и физические свойства липидного бислоя. Встраивание холестерина в фосфолипидный бислой вызывает как нарушение квазикристаллической упаковки цепей, так и уменьшение подвижности цепей – Эти эффекты холестерина называют, соответственно, «разжижающим» и «конденсирующим». При температуре, превышающей точку фазового перехода фосфолипида, холестерин уменьшает подвижность углеводородных цепей. При добавлении холестерина площадь молекулы лецитина уменьшается с 0,96 до 0,56 нм. Вот почему высокое содержание холестерина характерно для миелина и плазматических мембран, тогда как внутриклеточные мембраны содержат его в небольших количествах. В плотных миелиновых мембранах фосфолипиды и холестерин содержатся в отношении 1:1, а в менее плотных мито-хондриальных мембранах это отношение равно 3:1 или 8:1. Этот уплотняющий эффект холестерина максимален в районе центрального участка жирнокислотных радикалов и ослабевает в направлении концевых метальных групп. При температуре ниже точки фазового перехода фосфолипидов холестерин разжижает углеводородную область бислоя.
Фазовые переходы липидов при постоянной температуре могут быть вызваны изменениями заряда полярных групп липидов, возникающими при изменениях рН, ионной силы, концентрации ионов. Доказано, что температура фазового перехода есть функция величина заряда и плотности заряда на липидной молекуле. Любое увеличение заряда полярных групп благоприятствует жидкому состоянию из-за латерального электростатического отталкивания, тогда как уменьшение заряда обусловливает переход в твердокристаллическое состояние.
Важным путем изменения поверхностного заряда липидов в физиологических условиях является адсорбция катионов. Связывание катионов заряженными липидами сильно зависит от поверхностного потенциала, значительно различающегося в твердом и жидком состояниях из-за различий в молекулярной упаковке.
Освобождение или адсорбция катионов на мембранной поверхности может запускать фазовые переходы липидов. При определенных физиологических условиях структурные изменения липидов могут вызывать освобождение двухвалентных катионов с поверхности мембраны. Так, при переходе гель – жидкий кристалл с липидной поверхности освобождаются ионы кальция. Са+ 'и Mg+ стабилизируют организованную структуру, увеличивая температуру фазового перехода, а одновалентные катионы оказывают противоположный эффект. Двухвалентные катионы благоприятствуют гелеобразному, а одновалентные – жидкому состоянию мембраны. Поверхность липидов может рассматриваться как резервуар катионов, который способен регулироваться структурными изменениями.
■ Подводя итог вышеизложенному, можно заключить, что в организации липидов, в их асимметричном размещении, подвижности, модификации внутримолекулярных взаимодействий сокрыты многообразные регулирующие возможности.
Похожие работы
... в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой. Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение ...
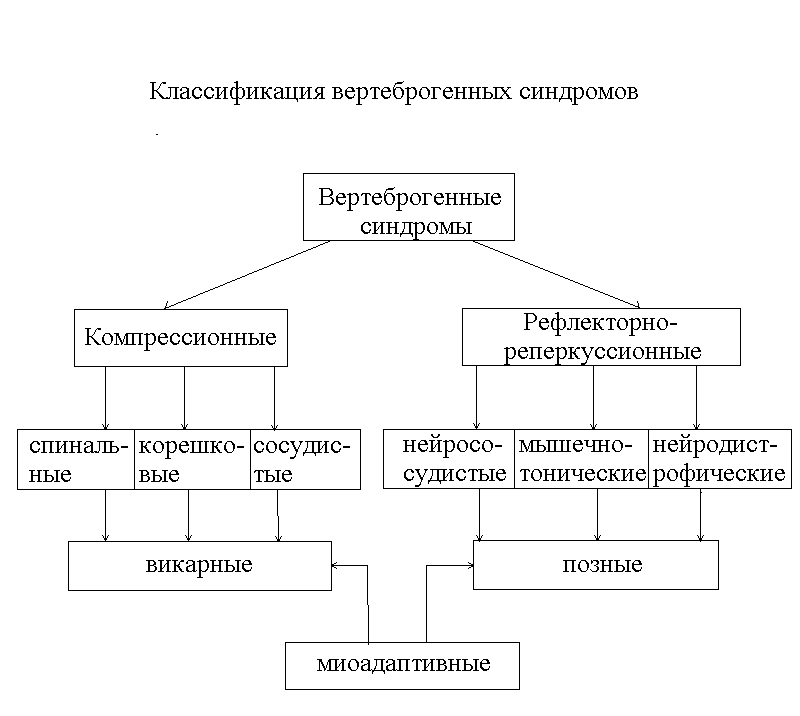
... , лептоспироз и др.) и вторичными (вертеброгенные, после детских экзантемных инфекций, инфекционного мононуклеоза, при узелковом периартериите, ревматизме и др.). По патогенезу и патоморфологии заболевания периферической нервной системы подразделяются на невриты (радикулиты), невропатии (радикулопатии) и невралгии. Невриты (радикулиты) – воспаление периферических нервов и корешков. По характеру ...
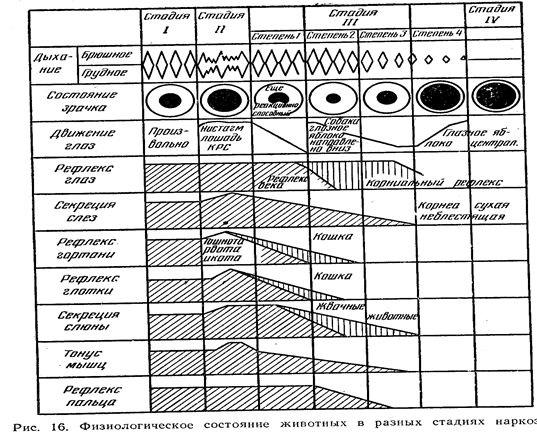

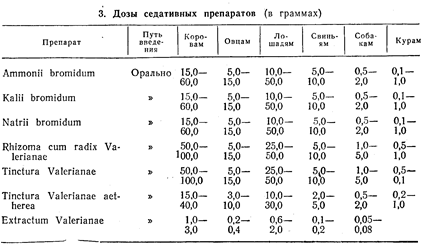

... , обволакивающие, адсорбирующие и др.). В этих же направлениях влияют местноанестезирующие вещества, выключающие отдельные звенья чувствительных нервов. Подавить боль можно, применяя вещества, угнетающие центральную нервную систему. Все эти средства используют при соответствующих показаниях. Но при болевых синдромах, являющихся следствием воспалительных процессов в организме, исключительно ...
... составляют тела клеток, дендриты и немиелинизированные аксоны, организованные в комплексы, которые включают бесчисленное множество синапсов и служат центрами обработки информации, обеспечивая многие функции нервной системы. Белое вещество состоит из миелинизированных и немиелинизированных аксонов, выполняющих роль проводников, передающих импульсы из одного центра в другой. В состав серого и белого ...
0 комментариев