РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
РОЛЬ БЕЛКОВ В ДИНАМИЧНОСТИ ЛИПИДНОГО БИСЛОЯ
МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
ЛИПИДЫ ВНЕШНЕЙ ЗОНЫ МЕМБРАН МОЗГА
Локализация ганглиозидов в головном мозге
Участие ганглиозидов в дифференциации клеток
Межклеточное гликозирование ганглиозидов
О-ацетилирование ганглиозидов – один из возможных механизмов изменения их структуры
ИЗМЕНЕНИЕ СОСТАВА ЛИПИДОВ В ОНТОГЕНЕЗЕ
Навигация
ИЗМЕНЕНИЕ СОСТАВА ЛИПИДОВ В ОНТОГЕНЕЗЕ
Липиды центральной нервной системы и структура клеточных мембран
82146
знаков
8
таблиц
16
изображений
9. ИЗМЕНЕНИЕ СОСТАВА ЛИПИДОВ В ОНТОГЕНЕЗЕ
Наиболее быстрое увеличение содержания липидов мозга наблюдается после периода интенсивного синтеза ДНК и белка, т.е. в период, когда происходит рост нейронов, глиальный митоз, аксодендритная пролиферация, формирование синаптических связей и, наконец, миелинизация.
До миелинизации липидный состав мозга сходен с другими органами, но миелинизация драматически изменяет состав липидов мозга. Правда, даже после завершения миелинизации содержание общих липидов в мозге человека продолжает увеличиваться до 30 лет и только после этого начинается их медленное снижение. Причем это снижение касается прежде всего фосфолипидов и жирных кислот и едва ощутимо затрагивает содержание холестерина и цереброзидов.
Липиды развивающегося мозга подразделяют на 4 группы на основе преимущественных: изменений в период миелинизации. Рассмотрим это на примере мозга крысы как объекта наиболее изученного, у которого лостнатальная миелинизация наиболее выражена в период с 21-го по 40-й день.
Содержание основных ганглиозидов мозга человека
| Ганглиозиды | Серое вещество | Белое вещество | ||
| новорожденные | взрослые | новорожденные | взрослые | |
| GM3 | 1 | - | 1 | |
| GM2 | 3,6 | 1,7 | 6,9 | 1>9 |
| GM1 | 14,6 | 12,8 | 19,1 | 12,6 |
| GDU | 71,6 | 22,8 | 57,8 | 18,4 |
| GDlb | 1,8 | 23,5 | 2,1 | 30,4 |
| GTI | 7,3 | 31,2 | 3,4 | 27,9 |
Первая группа липидов – эфиры холестерина и ганглиозиды. Концентрация их резко меняется в первые 6 дней постнатального развития крыс. Содержание эфиров холестерина уменьшается от 2 мкмолей на 1 г сырой массы до концентрации, составляющей менее 5% от начальной. У крыс это снижение происходит задолго до начала миелинизации, что отражает пролиферацию клеток или очень раннюю дифференциацию гли-альных клеток.
Ганглиозиды на 3-й день постнатального развития составляют 27% от содержания во взрослом организме. Концентрация ганглиозидов за 24 последующих дня быстро увеличивается, достигая 90% от уровня взрослого животного. Спектр индивидуальных ганглиозидов также меняется: при рождении преобладает моносиалоганглиозид G^m, а затем увеличивается содержание дисиалоганглиозидов. Увеличение количества ганглиозидов и изменение их состава связано с ростом аксонов и ден-дритов.
Вторая группа включает цереброзиды, сульфатиды, сфинго-миелин, трифосфоинозитиды, фосфатидные кислоты, галакто-зилдиглицериды. На 3-й день постнатального развития их концентрация невелика, а затем резко увеличивается в период от 12-го до 18-го дня. Пять первых перечисленных липидов являются основными комлонентами миелиновых мембран, их низкая концентрация при рождении подтверждает, что они локализованы в специальных мембранных структурах, которые появляются в мозге во время миелинизации. Полифосфоинозитиды и фосфатидные кислоты отличаются от других липидов этой группы, так как они продолжают заметно увеличиваться и после 24 дней, когда уровень других липидов этой группы стабилизируется.
Третья группа липидов включает фосфатидальэтаноламин, фосфатидальхолин, холестерин, фосфатидилсерин, фосфати-дилглицерин, концентрация которых составляет 12–34% от уровня взрослого организма и увеличивается во время миелинизации, но не столь значительно, как у липидов второй группы. Первые три представителя этой группы локализованы в мембранах миелина и нарастание их связано с миелинизацией.
Четвертая группа липидов охватывает три лииида мозга – фосфатидилэтаноламин, фосфатидилхолин и моно-фосфоинозитид, концентрация которых составляет 50–59% от содержания взрослого мозга и очень медленно увеличивается в период развития. Известно, что эти липиды являются повсеместными компонентами большинства мембранных структур и спектр их изменений не связан с преимущественными изменениями каких-либо специфических мембранных образований. Но в ходе онтогенеза в мембранах мозга увеличивается отношение ФЭ:ФХ и количество сфингомиелина. Диацильные формы фосфолипидов заменяются на плазмалогенные и значительно увеличивается микровязкость мембран.
Таким образом, различные классы липидов характеризуются индивидуальным характером накопления в период созревания мозга.
Выводы
1. Для нервной ткани характерно особенно высокое содержание липидов – до 50% от сухой массы ткани. Наряду с этим установлено огромное разнообразие и наличие специфических только для мозга индивидуальных липидов.
2. Фосфолипиды нервной ткани составляют до 70% от суммарного содержания липидов в сером веществе и до 45–50% – в белом веществе мозга. Обнаружена необычайно высокая гетерогенность фосфолипидов мозга по сравнению с висцеральными органами.
3. Основной представитель стеролов в нервной ткани – холестерин, на долю которого приходится около 25% от суммарного содержания липидов. В то же время в мозге взрослых животных мало эфиров холестерина.
4. Значительная часть сфинголипидов мозга представлена га-лактоцереброзидами и галактосульфатидами, количество которых в белом веществе значительно выше, чем в сером. Для мозга характерна высокая концентрация и большое разнообразие индивидуальных ганглиозидов.
5. Уровень свободных жирных кислот в мозге весьма невелик; напротив, установлено высокое содержание и огромное разнообразие жирных кислот в липидах нервной ткани. Основную массу жирных кислот липидов мозга составляют пальмитиновая 16:0, стеариновая 18:0, олеиновая 18:1 и арахидоновая 20:4 кислоты. В мозге идентифицировано около 40 индивидуальных жирных кислот, в том числе полиненасыщенных, длин-ноцепочечных и гидрокислот, которыми особенно богаты це-реброзиды и сульфатиды. Гетерогенность жирных кислот липидов мозга лежит в основе структурной лабильности мембран и определяет их важнейшие физико-химические свойства.
6. Содержание и соотношение отдельных классов липидов значительно изменяются в ходе развития и дифференцировки мозга. Наиболее интенсивно эти процессы протекают в раннем постнатальном онтогенезе.
7. Установлены существенные различия в липидном составе важнейших мембранных образований нервной ткани. Обращает на себя внимание высокое содержание и чрезвычайное разнообразие ганглиозидов, особенно в мембранах нервных окончаний и в дендритах. Именно здесь наиболее полно проявляется функциональная роль этих специфических липидов, участвующих в связывании различных катионов, в процессах адгезии, в обеспечении иммунохимической специфичности и др.
8. Специфическими липидными компонентами миелина являются цереброзиды и сульфоцереброзиды; установлено высокое содержание в миелине холестерина и фосфолипидов, в том числе плазмалогенов, доля которых в миелине составляет более 90% от его количества в целом мозге.
9. Липиды мембран мозга организованы в бислой с планар-ной и поперечной асимметричностью размещения липидов по слоям. Она поддерживается механизмами, учитывающими структуру липидов, их ненасыщенность, стереоконфигурацию полярных групп, специфичность липид-переносящих белков, ферментативные превращения липидов.
10. Динамичность липидного бислоя определяется интрамолекулярными движениями и фазовыми переходами липидов, что создает основу для структурных перестроек в мембранах.
11. Липиды бислоя принимают участие в передаче информации через мембрану и в осуществлении внутриклеточного ответа.
12. Организованная многослойная структура миелина, имеющая самое высокое содержание липидов, поддерживается длинно- и короткорадиусными взаимодействиями между липидами и основным и протеолипидным белками. Формирование миелина является сложным синхронизированным процессом взаимодействия аксонаиглии, любое его нарушение вызывает демиелинизацию.
13. Экстраклеточный матрикс мембран мозга представляет собой комплексную, динамичную систему, где происходит распределение регуляторных сигналов, передача информации внутрь клетки, реализуется прямая и обратная связь с ядром.
14. Специфичность экстраклеточного матрикса определяется первичной структурой, организацией и площадью, занимаемой гликолипидами и гликопротеинами. Экстраклеточный матрикс нейрональных мембран обогащен разнообразными ганглиозидами.
Похожие работы
... в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой. Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение ...
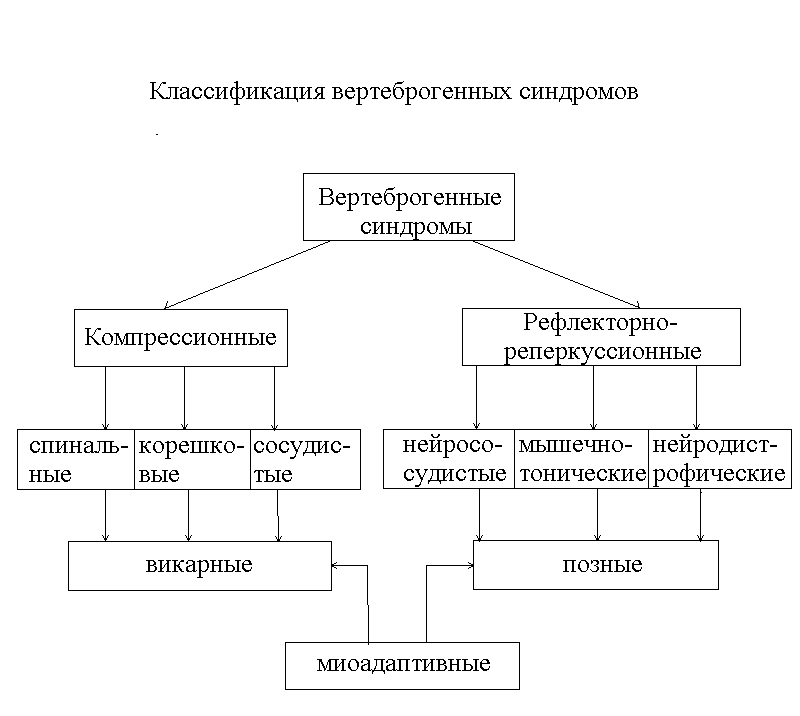
... , лептоспироз и др.) и вторичными (вертеброгенные, после детских экзантемных инфекций, инфекционного мононуклеоза, при узелковом периартериите, ревматизме и др.). По патогенезу и патоморфологии заболевания периферической нервной системы подразделяются на невриты (радикулиты), невропатии (радикулопатии) и невралгии. Невриты (радикулиты) – воспаление периферических нервов и корешков. По характеру ...
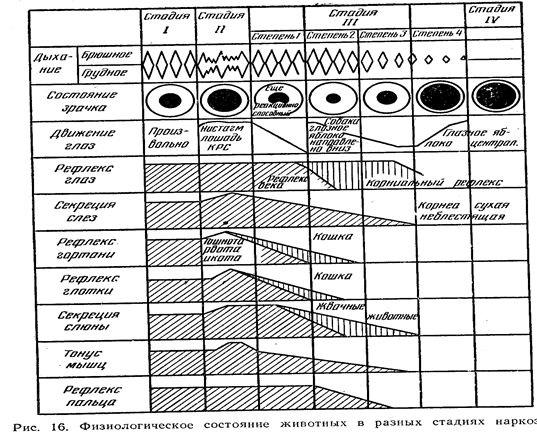

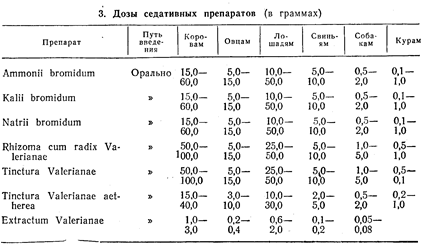

... , обволакивающие, адсорбирующие и др.). В этих же направлениях влияют местноанестезирующие вещества, выключающие отдельные звенья чувствительных нервов. Подавить боль можно, применяя вещества, угнетающие центральную нервную систему. Все эти средства используют при соответствующих показаниях. Но при болевых синдромах, являющихся следствием воспалительных процессов в организме, исключительно ...
... составляют тела клеток, дендриты и немиелинизированные аксоны, организованные в комплексы, которые включают бесчисленное множество синапсов и служат центрами обработки информации, обеспечивая многие функции нервной системы. Белое вещество состоит из миелинизированных и немиелинизированных аксонов, выполняющих роль проводников, передающих импульсы из одного центра в другой. В состав серого и белого ...
0 комментариев