РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
ДИНАМИЧНОСТЬ БИЛИПИДНОГО СЛОЯ МЕМБРАНЫ
РОЛЬ БЕЛКОВ В ДИНАМИЧНОСТИ ЛИПИДНОГО БИСЛОЯ
МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
ЛИПИДЫ ВНЕШНЕЙ ЗОНЫ МЕМБРАН МОЗГА
Локализация ганглиозидов в головном мозге
Участие ганглиозидов в дифференциации клеток
Межклеточное гликозирование ганглиозидов
О-ацетилирование ганглиозидов – один из возможных механизмов изменения их структуры
ИЗМЕНЕНИЕ СОСТАВА ЛИПИДОВ В ОНТОГЕНЕЗЕ
Навигация
Участие ганглиозидов в дифференциации клеток
Липиды центральной нервной системы и структура клеточных мембран
82146
знаков
8
таблиц
16
изображений
8.4 Участие ганглиозидов в дифференциации клеток
Была предложена модель клеточного цикла, в которой кроме стадии покоя G0, неустойчивой и регулируемой циклическими нуклеотидами, постулируется стадия D-дифференциации, контролируемая ганглиозидами. По мере формирования ней-рон-нейрональных взаимодействий меняется структура и количество ганглиозидов и увеличивается число высокоаффинных контактов.
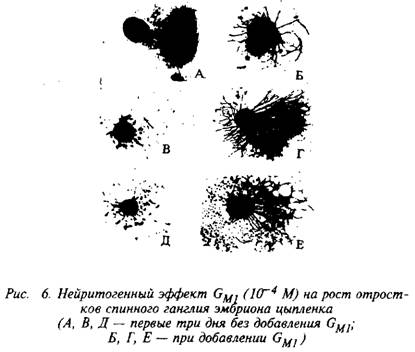
Участвуя в дифференциации клеток, ганглиозиды увеличивают время выживания клеток и вызывают морфологические изменения клеток, проявляя нейритогенный эффект. Нейритогенный эффект экзогенных ганглиозидов обнаружен в культурах клеточных линий нейронального и хромаффинного происхождения, в симпатических и парасимпатических ганглиях и в нервно-мышечных препаратах.
На рис. 6 представлен нейритогенный эффект моносиалоганглиозида GMI в концентрации 10~М на рост отростков спи-нального ганглия эмбриона цыпленка. Экзогенные ганглиозиды оказывают влияние на протяженность отростков, их число на клетку и на разветвленность отростков. Интересно, что моносиалоганглиозид GM1 вызывает только увеличение длины аксонов, а три- и тетрасиалоганглиозиды в тех же концентрациях усиливают спрутинг и арборизацию.
8.5 Терапевтические эффекты ганглиозидов
Ганглиозиды in vivo обладают уникальными свойствами: при введении в организм подкожно, внутримышечно или интраперитонеально они относительно длительное время сохраняются в кровяном русле, лишены токсичности, в небольших количествах проникают через гемато-энцефалический барьер и активно встраиваются в нейрональные мембраны. Они способствуют репарации поврежденных аксонов, обладают выраженными терапевтическими эффектами при травмах головного и спинного мозга.
В настоящее время наиболее изучена молекулярная и биологическая роль в этих процессах моносиалоганглиозида GM1, который при введении in vivo:
а) восстанавливает нейрохимические параметры дофаминер-гических нейронов после нарушения нигростриатной системы, усиливает захват дофамина и активность ирозингидроксилазы;
б) восстанавливает нейрохимические характеристики при частичной холинергической и глутаматергической деафферен-тации гиппокампа, увеличивает активность холинацетилтрансферазы и ацетилхолинэстеразы;
в) восстанавливает высокоаффинный захват холина в коре больших полушарий после нарушений ядер переднего мозга;
г) нормализует дисбаланс между активностью дофамин- и серотонинергических нейронов, вызванный введением апомор-фина;
д) оказывает рост-стимулирующий эффект и защитное действие против вторичной дегенерации серотонин- и норадре-нергических нейронов, вызванной нейротоксинами;
е) уменьшает церебральный отек и восстанавливает ионный баланс после травмы;
ж) способствует регенерации зрительного нерва после перерезки.
С другой стороны, введение антител к GM] вызывает у развивающихся животных нарушение дендритной арборизации и поведения, ухудшение обучаемости, появление эпилептиформ-ной активности.
Моносиалоганглиозид GM1 хорошо внедряется в мембраны, причем особенно хорошо встраивается молекула GMI, имеющая в своем составе С 2о-эритросфингозин. Возможно, это объясняется его более высокой способностью к мицеллообразованию. Он образует дископодобные мицеллы с М^ЗОО кД, имеющие гидродинамический диаметр около 60 нм.
Интересно, что мицеллы из моносиалоганглиозида GMI по-тенциируют действие ионофора грамицидина D. Ганглиозид-ные мицеллы с заключенными в них молекулами грамицидина включаются в модельную мембрану из фосфатидилсерина и изменяют ее проводимость для ионов калия. После добавления мицелл с ионофором увеличивается время открытия ионных каналов и изменяется их амплитуда.
■ Таким образом, ганглиозидные мицеллы могут участвовать в ионтранспортном процессе в мембране, «маркируя» входы в селективные ионные каналы,
О механизмах и функциональной последовательности действия ганглиозидов известно мало. Встраивание экзогенных ганглиозидов, приводящее к перестройке мембранных ансамблей, изменяет ряд внутриклеточных процессов. Вызванная ганглиозидами дифференциация сопровождается изменением активности Na5K-ATOa3bi, увеличением внутриклеточного уровня цАМФ, уменьшением включения меченого тимидина в ДНК и значительным удлинением фазы G( клеточного цикла. Внедрение ганглиозидов вызывает немедленную перестройку мик-рофиламентной и микротубулиновой системы клеток.
Включение в мембрану экзогенных ганглиозидов усиливает аксональный ток гликозилированных белков и липидов, увеличивает количество гликопротеинов с терминальной манно-зой. Внедрение моносиалоганглиозида GM1 увеличивает в мембране количество эндогенных моносиалоганглиозидов и изменяет активность гликозилтрансфераз: усиливается активность эктофукозилтрансферазы при неизменности активностей сиалил-и галактозилтрансфераз. Внедрение в мембрану трисиалоганг-лиозида GTlb вызывает противоположный эффект.
Недавно выявлено влияние индивидуальных ганглиозидов на фосфорилирование гистона Hj и тубулина, причем в отношении фосфорилирования гистона были особенно эффективны GOJb >GDl* >GTia >GD3> а тубулина – Gxlb > GTla > GQlb> GDla. Показано, что тетрасиалоганглиозид Gglb проявляет зависимое от концентрации влияние на активность Са+ – фос-фолипид-, Са +-кальмодулин-, цАМФ- и цГМФ-активируемых протеиякиназ
Похожие работы
... в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой. Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение ...
... , лептоспироз и др.) и вторичными (вертеброгенные, после детских экзантемных инфекций, инфекционного мононуклеоза, при узелковом периартериите, ревматизме и др.). По патогенезу и патоморфологии заболевания периферической нервной системы подразделяются на невриты (радикулиты), невропатии (радикулопатии) и невралгии. Невриты (радикулиты) – воспаление периферических нервов и корешков. По характеру ...

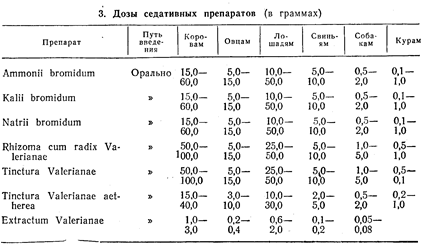

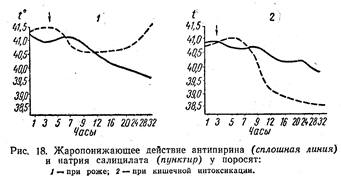
... , обволакивающие, адсорбирующие и др.). В этих же направлениях влияют местноанестезирующие вещества, выключающие отдельные звенья чувствительных нервов. Подавить боль можно, применяя вещества, угнетающие центральную нервную систему. Все эти средства используют при соответствующих показаниях. Но при болевых синдромах, являющихся следствием воспалительных процессов в организме, исключительно ...
... составляют тела клеток, дендриты и немиелинизированные аксоны, организованные в комплексы, которые включают бесчисленное множество синапсов и служат центрами обработки информации, обеспечивая многие функции нервной системы. Белое вещество состоит из миелинизированных и немиелинизированных аксонов, выполняющих роль проводников, передающих импульсы из одного центра в другой. В состав серого и белого ...
0 комментариев