Літературний огляд
Біохімія та мікробіологія процесу
Умови життєдіяльності дріжджів
Види та джерела харчування
Біохімія бродіння та дихання
Мікроорганізми – супутники дріжджів
Технологічна частина
Асортимент і характеристика готової продукції
Характеристика сировини та допоміжних матеріалів
Розрахунок основного та допоміжного обладнання
Мм – товщина днища
Тепловий баланс ректифікаційної колони
Розрахунок та підбір допоміжного обладнання
Економічна частина
Розрахунок чисельності виробничого персоналу та фонду заробітної плати
Розрахунок ефективності заходу
Організація управління охороною праці на підприємстві
Перелік шкідливих і небезпечних виробничих чинників
Освітлення
Пожежна безпека
Навигация
Види та джерела харчування
Технологія виробництва спирту на спиртовому заводі "Nemiroff"
132766
знаков
23
таблицы
2
изображения
2.2.3 Види та джерела харчування
Розрізняють екзогенне та ендогенне харчування дріжджів: при екзогенному харчуванні поживні речовини потрапляють до клітини із зовнішнього середовища, при ендогенному дріжджі використовують (в основному при голоданні) свої резервні речовини: глікоген, трегалозу, ліпіди азотисті сполуки.
Вуглецеве харчування. Дріжджі (Saccharomyes cerevisiae) використовують вуглець із різноманітних органічних сполук: глюкози, маннози, галактози, фруктози (D - форми). Пентози Sacch. cerevisiae не асимілюють. При відсутності гексоз джерелом вуглецю можуть бути також гліцерин, маніт, етиловий та інші спирти, органічні кислоти (молочна, оцтова, яблучна, лимонна).
Слід враховувати поліауксію – послідовність споживання різноманітних джерел вуглецю. При періодичному культивуванні в першу чергу споживаються глюкоза та фруктоза. Послідовність засвоювання жирних кислот залежить від раси дріжджів, що використовуються, та складу цих кислот. Наприклад, оцтова кислота перешкоджає споживанню молочної, а молочна – гліколевий. Оцтова кислота і глюкоза засвоюються одночасно. Як правило, в першу чергу засвоюється з суміші те джерело вуглецю, яке забезпечує найбільшу швидкість росту дріжджів.
При безперервному культивуванні дріжджів зі збільшенням швидкості розбавлення середовища в ньому залишається більше вуглецевого компоненту, який засвоюється останнім.
Дісахариди, з яких спиртові дріжджі використовують мальтозу та сахарозу, попередньо підпадають під вплив гідролізу відповідними ферментами дріжджів до моносахаридів. При переході від анаеробних умов до аеробних послаблюється можливість дріжджів зброджувати глюкозу та мальтозу, а сахарозна активність їх підвищується в 2,5 рази. Дріжджі споживають мальтозу тільки при відсутності в середовищі фруктози та глюкози. Мальтоза зброджується майже повністю під час стаціонарної фази росту дріжджів.
Органічні кислоти мають важливе значення при метаболізмі вуглецю, енергетичному обміні мікроорганізмів, синтетичних та дисиміляційних процесах. Використання кислот жирного ряду в якості джерела вуглецю залежить від виду та раси дріжджів, концентрації кислоти, довжини її вуглецевого ланцюга та ступеня електролітичної дисоціації. Непоганими субстратами служать кислоти з довжиною вуглецевого ланцюга від С2 до С4 при порівняно низькій концентрації. Калійні солі кислот, що містять у молекулі від 2 до 5 атомів вуглецю, стимулюють ріст дріжджів в 1,4 – 3,3 рази сильніше порівняно з відповідними кислотами.
Жирні кислоти з середньою довжиною вуглецевого ланцюга (від С6 до С10) в значно меншій мірі споживаються дріжджами і тільки в умовах дуже низької концентрації (0,02 – 0,05%). При більш високій концентрації розвиток дріжджів пригнічується. Жирні кислоти з 12 – 17 атомами вуглецю в молекулі засвоюються вибірково в залежності від роду та виду дріжджів.
Будь-який з проміжних продуктів циклу Кребса (піровиноградна, лимонна, янтарна, фумарола, яблучна кислоти) можуть бути єдиними джерелами вуглецю для життєдіяльності дріжджів.
Азотне харчування. Дріжджі мають можливість синтезувати всі амінокислоти, що входять до складу їх білка, безпосередньо з неорганічних азотистих сполук при використанні в якості джерела вуглецю органічних сполук - проміжні продукти розпаду вуглеводів, які утворюються при диханні та бродінні.
Дріжджі Sacch. Cerevisiae засвоюють лишень дві форми азоту: аміачний та органічних речовин. Ці мікроорганізми ефективно використовують азот сульфату і фосфату амонію, мочевини, аміачних солей оцтової, молочної, яблучної та янтарної кислот. В присутності цукрів, що зброджуються. Аміачні солі є для дріжджів джерелом лише азоту; однак при використанні його звільнюються кислоти, що змінюють рН середовища. Аміачний азот використовується дріжджами краще, ніж азот багатьох амінокислот.
Амінокислоти – одночасно джерело азоту та вуглецю, при цьому останній засвоюється з кетокислот, що утворюються в результаті відщеплення аміногруп. Можлива і безпосередня асиміляція амінокислот з поживного середовища, яке містить їх повний набір та будь-який цукор, що зброджується. Як наслідок знижуються витрати цукру середовища на харчування дріжджів та дещо збільшується вихід спирту при бродінні.
Завдяки асиміляції амінокислот забезпечується синтез білка, в тому ж числі і ферментів, активуються деякі ферменти, що вже є в клітині, прискорюється процес брунькування дріжджових клітин.
Для споживання органічного азоту (амінокислот, амідов) багатьом дріжджам необхідні вітаміни (біотин, пантотеновая кислота, тіамін, пірідоксін та ін.). дріжджі не засвоюють такі азотисті сполуки, як білки, бетаїн, холін, пурини та аміни у вигляді етіламіна, пропіл- та бутіламіна. Пептиди займають середнє положення між амінокислотами та білками. Споживання дріжджами пептидів знижується з підвищенням їх складності. Деяка кількість пептидів в середовищі поряд з іншими формами азоту сприяють використанню амінокислот.
На утворення 10 млрд. дріжджових клітин витрати азоту в умовах анаеробіозу складають 66 – 77 мг, в умовах аеробіозу 37 -53 мг. Про умови культивування та фізіологічний стан дріжджів говорять по вмісту азоту в них, який залежить від складу середовища, кількості додатково вводимих поживних речовин та від раси дріжджів. В дріжджах, що отримують на спиртових заводах, загального азоту 7 – 10% (іноді до 12) на суху речовину.
Фосфорне харчування. В анаеробних умовах дріжджі засвоюють фосфор головним чином у початковому періоді бродіння – 80-90% від максимальної кількості в дріжджах. Молоді дріжджові клітини, що енергійно розмножуються багатші на фосфор у порівнянні зі старими, що не брунькуються. Наприклад, після 6 годин бродіння дріжджі містять 2,15% фосфору на суху речовину, до кінця бродіння – лише 1%.
2.2.4 Інші фактори
Швидкість росту дріжджів залежить від різниці осмотичного тиску в клітині та в суслі: чим вона більша, тим швидше розмножуються дріжджі. В наслідок цього більш активний фізіологічний стан дріжджів спостерігається при зброджуванні меляси за двох поточною схемою.
При обробці спиртових дріжджів ультразвуком в декілька разів підвищується активність інвертази, в деяких випадках стимулюється їх ріст.
Спирти, ефіри та слабкі розчини луг розчиняють липоїдниї речовини клітин дріжджів. спирти навіть у невеликих концентраціях (3 – 4%) гальмують брунькування дріжджів. однак в безперервному потоці середовища, що зброджується, дріжджі могуть розмножуватися при відносно високій концентрації спирту (7 – 8 об. %) та продовжувати зброджування цукрів до концентрації 10 – 12 об. %. розмноження дріжджів при безперервному зброджуванні залежить головним чином від вмісту поживних речовин та менше від кількості спирту в середовищі.
Формалін, кислоти та солі важких металів відносяться до плазматичних отрут. Невелика кількість формаліна (0,09%) порушує нормальну життєдіяльність дріжджів, а доза 0,001% гальмує їх брунькування. Зазвичай дози речовин, знижують бродильну енергію дріжджів, значно вище доз, затримуючих брунькування.
Сірчаниста, азотиста та фтористоводородна кислоти й їх солі в дуже малих концентраціях перешкоджають нормальному росту дріжджів.
Вільні органічні кислоти оказують більш інгібіруючу дію на дріжджі, ніж їх солі. Летючі органічні кислоти навіть в незначних концентраціях пригнічують їх розмноження та прискорюють їх відмирання. Найбільш сильні інгібітори – мелясна та капронова кислоти. Особливо чуттєві дріжджі до летючих органічних кислот при зниженні рН середовища до 4. в цих умовах через добу в дріжджовій популяції спостерігається велика кількість плазмолізованних клітин та бруньок.
Мурашина кислота знижує коефіцієнт розмноження дріжджів, не викликаючи при цьому відмирання клітин. Оцтова кислота - порівняно слабкий інгібітор.
Деякі важкі метали в дуже малих концентраціях вбивають дріжджові клітини (срібло – 0,000001%, мідь – 0,005%), а в концентраціях, що не піддаються визначенню хімічним аналізом, гальмують ріст дріжджів. Бактерицидна дія важких металів залежить від складу середовища, її кислотності, температури та густини дріжджової популяції.
В разі присутності фурфуролу в середовищі, що зброджується, зменшується кількість клітин, що брунькуються, та їх розмір. Навіть при незначному вмісті фурфуролу знижується мальтозна і зимазна активність дріжджів, що були виділені з мелясної бражки.
Сульфонал у невеликих концентраціях (70 – 100 г на 1 т меляси) не впливає на життєдіяльність дріжджів і пригнічує молочнокислу мікрофлору. Хлор, хлорне вапно, манганокислий калій, сильно окислюючи органічні сполуки, руйнують їх.
В бражках з підвищеним вмістом йонів Са, Мg, Fe у дріжджових клітин втрачається водна оболонка, через що зменшується йонна сфера й електричний заряд на поверхні клітин і створюються умови для аглютинації дріжджів.
Спиртові раси дріжджів мають негативний електрокінетичний потенціал: від -7 до -13 мВ, через що вони адсорбують на своїй поверхні меланоїдини з позитивним потенціалом. Зі зниженням рН середовища електрокінетичний потенціал меланоїдинів зростає, у зв’язку з чим збільшується ступінь адсорбції їх на дріжджових клітинах. Меланоїдини присвоюють дріжджам темний колір, сприяють відмиранню дріжджових клітин, і, як наслідок, приводять до зниження їх ферментативної активності, а саме активності інвертази та каталази.
Десорбція фарбуючи речовин з поверхні дріжджової клітини проходить інтенсивно при рН промивної води вище 9. При рН близько 3 фарбуючи речовини не десорбуються.
Багато ферментів дріжджів активуються в присутності сульфгідрильних сполук, що містять SH-групи, таких, як цистеїн, глютатіон. Ці сполуки легко перетворюються одне в друге, мають важливе значення в активуванні та шелюгуванні дії багатьох окислювально-відновлювальних і гідролітичних ферментів, що визначають життєдіяльність та обмінні процеси мікроорганізмів.
SH-групи грають важливу роль в ланцюгу окислювально-відновлювальних реакцій, і являються необхідною ланкою в передаванні електрона від сукцината до кисню повітря через цитохром. Активність багатьох дегідрогеназ, флавонових та пірідоксалевих ферментів пов’язана з наявністю в молекулі вільних SH-груп.
Відновлений глютатіон та цистеїн прискорюють спиртове зброджування внаслідок відновлення SH-групи толових ферментів, що беруть участь в анаеробному та аеробному окислюванні цукрів. Однак використання цих речовин дуже дорого коштує та економічно недоцільне; в якості їх аналогів може бути використаний дріжджовий автолізат.
Похожие работы
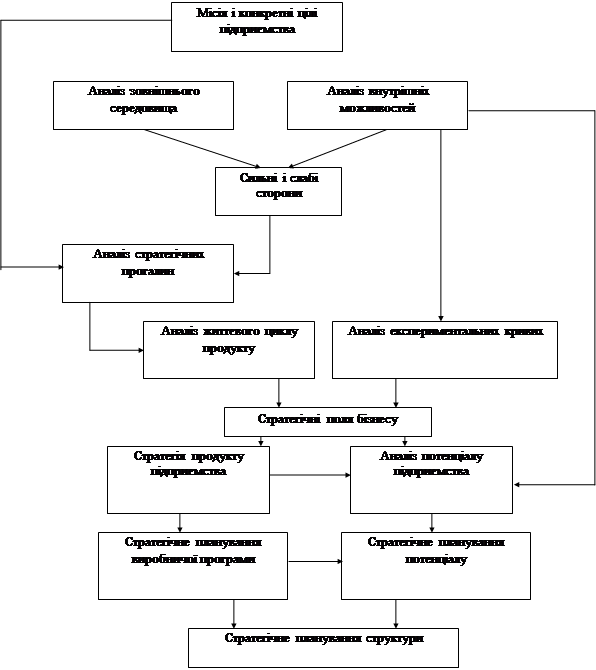
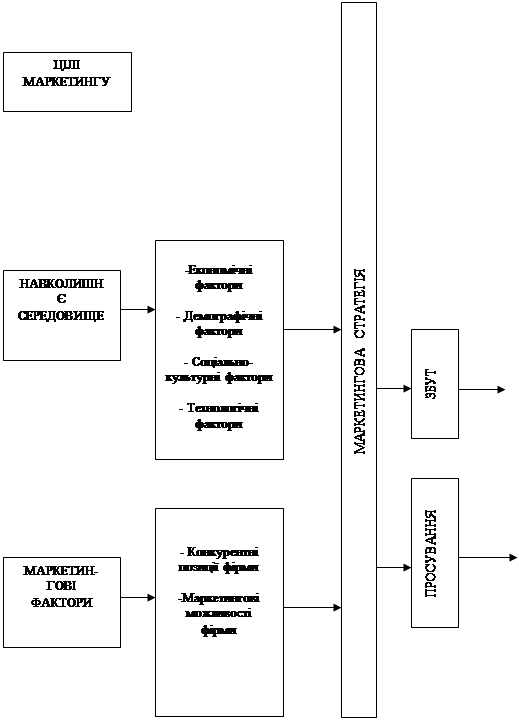
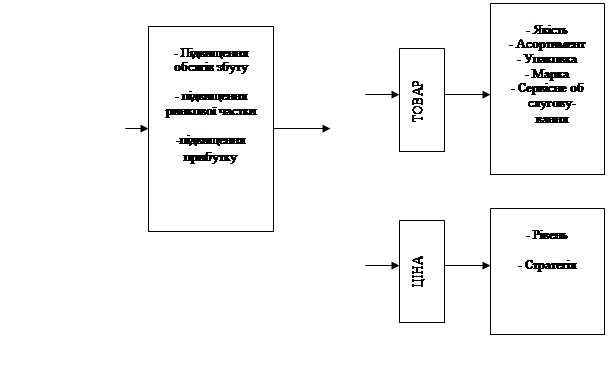
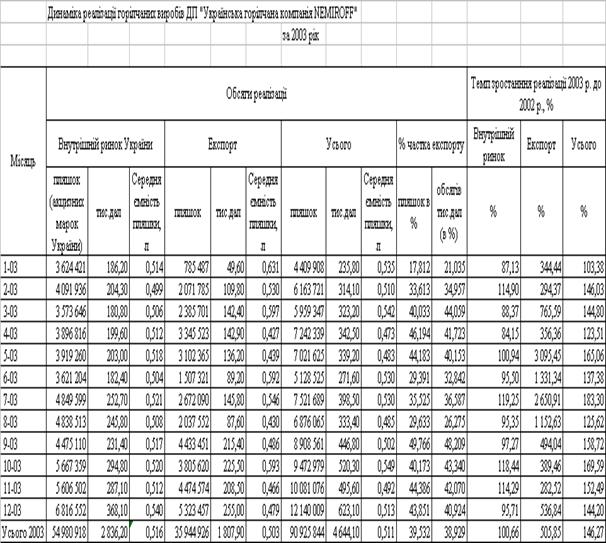
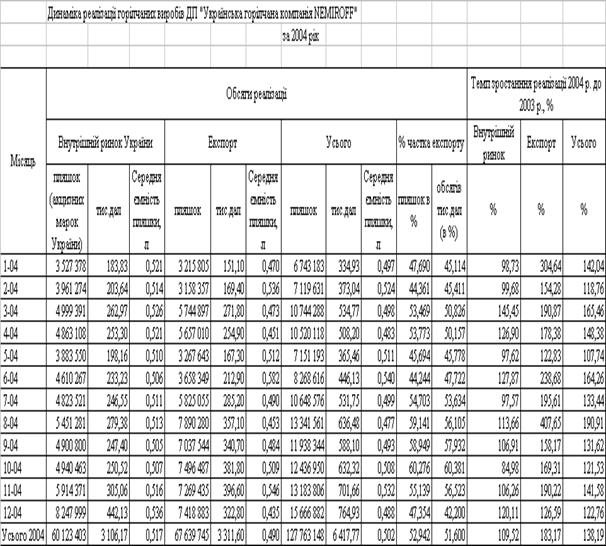
... Розділ 2. Аналіз зовнішньоекономічної діяльності дп “українська горілчана компанія nemiroff” та її положення на світовому ринку алкогольної продукції 2.1 Загальна характеристика діяльності ДП “Українська горілчана компанія NEMIROFF" Українська горілочна компанія Nemiroff - дочірнє підприємство кіпрської компанії Biostar Investments Ltd., на ринку працює з 1992 року [51]. У 2006 році в ...
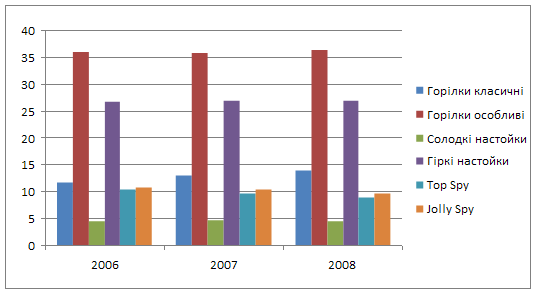
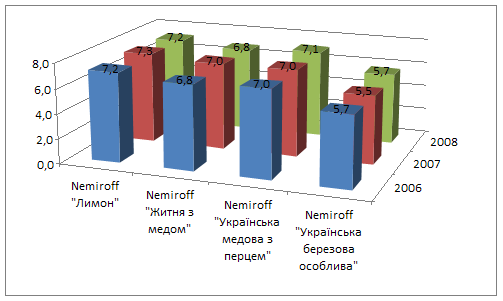
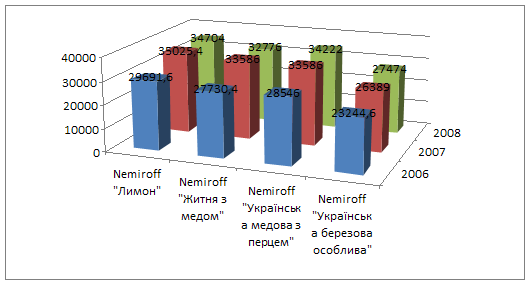
... у порівнянні з 2007 роком. зниження фондовіддачі і зростання фондоємності негативно вплинуло на діяльність підприємства. Рентабельність у 2008 році збільшилась на 6,54%, у порівнянні з 2006 роком та на 11, 94%, у порівнянні з 2007 роком. 2. Товарна номенклатура Продукція марки Nemiroff ― це відмінна якість за доступною ціною. Продукція заводу користується визнанням в багатьох регі ...
... час як обсяги імпорту горілки впали (зокрема, імпорт російської горілки в Україну у 2006 році, порівняно з 2005, скоротився за оцінками експертів, майже на 20%, а частка російської горілки на українському ринку не переважає 1%). За оцінками експертів, до 80% імпортної продукції на алкогольному ринку Росії – українського походження, а частка її на російському ринку в цілому у першому півріччі 2006 ...
... таким чином, щоб обмежити доступ імпортерів на місцеві ринки. Тим часом звуження присутності в Росії, що є другим споживачем алкоголю у світі, стане вагомою втратою для українців. За прогнозами аналітиків, російський ринок горілки протягом найближчих п'яти років щорічно зростатиме на 8,2% при тому, що торік його обсяг у роздрібних цінах становив $37,5 млрд. Минулоріч виробники української горі ...
0 комментариев