Щелочные протеиназы
рода Bacillus
Получение
щелочных протеиназ
Субстратная
специфичность
микробных липаз
Применение
микробных липаз
Методы исследования
Методика изучения
культуральных
свойств
Приготовление
бактериальной
суспензии
Определение
протеолитической
активности
(метод Вильштеттера
и Вальдшмидт
– Лейтца в
модификации)
Качественный
анализ вещества
по ИК-спектрам
поглощения
Определение
токсичности
сточных вод
Изучение
морфолого-физиологических
и культуральных
свойств микроорганизмов
Влияние условий
культивирования
на свойства
ферментного
препарат а,
продуцируемого
культурой рода
Listeria
Изучение возможности
проведения
процесса
обезжиривания
I с применением
ферментного
препарата
Экономическая
часть
Безопасность
жизнедеятельности
Требования
к микробиологическим
лабораториям
Охрана окружающей
среды. спав и
их вовлечение
в метаболические
процессы
Навигация
Щелочные протеиназы рода Bacillus
Выделение, изучение свойств микроорганизмов и их использование для выполнения подготовительных процессов переработки овчинно-мехового сырья
198182
знака
14
таблиц
0
изображений
1.2 Щелочные протеиназы рода Bacillus
Внеклеточные щелочные протеиназы выполняют ряд важных катаболических функций вне клетки. Наиболее очевидной функцией щелочных протеиназ является расщепление белков и других высокомолекулярных субстратов, содержащихся в питательной среде, и превращение их в форму, способную легко проникать внутрь микробной клетки. Щелочные протеиназы играют определенную роль в других жизненно важных процессах клетки. Предполагается, что сериновая протеиназа выполняет три внутриклеточные функции: 1) участвует в синтезе белковой оболочки, вероятно, снабжает аминокислотами, 2) играет роль мусорщика – убирает ненужные вещества клетки, 3) принимает участие в модификации ферментов, особенно РНК-полимеразы /23/.
В 1938 г. Кунитц впервые высказал предположение о существовании связи между процессом спорообразования и активностью щелочной протеиназы. Внеклеточные щелочные протеиназы бактериального происхождения, главным образом из Bacillus subtilis, изучены более детально. Это – ферменты широкой специфичности, гидролизуют до 80 связей в белках, с оптимумом рН в щелочной зоне. Впервые фермент был выделен, очищен и охарактеризован в лаборатории Карлсберга (Копенгаген). Фермент был назван субтилопептидазой А, алкалазой, субтилизином А, а затем за ним закрепилось название субтилизин Карлсберг. Позднее, в той же лаборатории из другого штамма Bac. subtilis был получен другой фермент. Этот фермент получил название субтилизина Novo. Несмотря на большое сходство с субтилизином Карлсберг, он все же не был идентичен ему, отличаясь некоторыми физико-химическими и энзиматическими свойствами. Одновременно с этими исследованиями в Японии была выделена протеиназа, продуцируемая культурой Bac.amyloliquefaciens. Она получила название BPN’. Таким образом, в настоящее время изучено три щелочные протеиназы типа субтилизина. Они имеют много общих свойств, но тем не менее субтилизин Карлсберг отличается от субтилизинов Novo и BPN’, являющихся идентичными. Молекулярный вес субтилизинов 27 000. При рН 5,0 и ниже фермент быстро и необратимо инактивируется. Оптимум рН действия для всех субтилизинов лежит в области рН 9–10. Ферменты проявляют широкую специфичность при гидролизе белков. Обладают эстеразной активностью /24/.
Несмотря на значительное сходство: 1) в ферментативном поведении, 2) гидролизе одних и тех же субстратов, 3) сходной реакции по отношению к одним и тем же ингибиторам, 4) близости происхождения, субтилизины отличаются между собой. Изоэлектрическая точка субтилизина Карлсберг лежит в зоне рН 9,4, субтилизина Novo и BPN’ – 7,8. Субтилизин состоит из одной пептидной цепи и 275 аминокислотных остатков. Субтилизины – одни из немногих белков, для которых определна трехмерная структура и построена модель молекулы.
По механизму действия и субстратной специфичности субтилизины близки к сериновым протеиназам животного происхождения – химотрипсину, трипсину, несмотря на отсутствие какого бы то ни было сходства в структуре между ними /25/.
Характер регуляции биосинтеза щелочных протеиназ изучен недостаточно. Щелочным протеиназам принадлежит ряд важных функций в процессах клеточной дифференцировки, в частности спорообразования. Известно, что щелочные протеиназы различных типов образуются в основном спорообразующими видами бактерий, причем образование фермента тесно связано с процессом спорообразования /26/.
Спорообразующие виды бактерий являются продуцентами многих биологически активных соединений. Образование ряда антибиотиков, ферментов и токсинов начинается после завершения активного роста продуцентов и совпадает с началом их споруляции. Поэтому выяснение связи процесса споруляции с образованием различных биологически активных соединений и изучение функций этих соединений представляет важность для разработки как общих принципов, так и частных методов селекции высокопродуктивных штаммов спорообразующих микроорганизмов. Следует отметить, что большинство продуцентов, характеризующихся сверхсинтезом фермента, в генетической плане практически не изучены /27/.
Исследования подобного рода представляют также определенный интерес и в связи с изучением механизма дифференцировки спор, поскольку ряд биологически активных соединений, накапливающихся при споруляции, по-видимому, участвует в регуляции различных этапов этого процесса.
1.2.1 Генетика и физиология спорообразования различных видов рода Bacillus
Жизненный цикл спорообразующих бактерий состоит из прорастания споры, вегетативной стадии, перехода к споруляции и завершается образованием зрелой споры. Причем изменения структуры клетки при переходе в споруляционное состояние осуществляются во времени и составляют несколько этапов.
На модели Bac. subtilus установлена последовательность из семи этапов споруляции /28/, характеризующихся определенными морфологическими изменениями. Выяснению последовательности этапов спорообразования во много способствовало также выделение мутантов, неспособных образовывать зрелые споры – «spo» – мутанты. В зависимости от нарушения определенного этапа спорообразования данные мутанты классифицируют как spo0, spoI, spoII и та далее.
Генетическое нарушение определенного этапа спорообразования характеризуется отсутствием ряда морфологических и биохимических функций, т.е. определенными фенотипическими последствиями. Например, характерной чертой spo0-мутантов, имеющих нарушение «0» стадии споруляции, является плейотропность, т.е. наряду с нарушением способности клеток спорулировать не осуществляется ряд биохимических событий, таких, как образование протеиназ, пептидных антибиотиков, сохраняется чувствительность к некоторым фагам и исчезает компетентность при трансформации.
Более детальный анализ показал, что как переход от вегетативной стадии роста к спорообразованию, так и переключение с одного этапа спорообразования на другой коррелирует с рядом биохимических изменений в клетке, в частности со снижением синтеза общей РНК, модификацией синтеза ДНК, а также с появлением определенных биологически активных соединений. Результаты генетических исследований различных классов мутантов показали, что процесс спорообразования определяется полигенной системой, локализующейся в различных районах хромосомы Bac. subtilus (более 100 генов). В настоящее время на хромосоме Bac. subtilus локализовано 28 оперонов, отвечающих за VII этапов процесса спорообразования /29/.
Считается, что регуляция спорообразования может осуществляться на нескольких уровнях: на уровне транскрипции, трансляции и посттрансляционной регуляции.
Регуляция спорообразования на уровне транскрипции. Результаты исследования перехода клеток Bac. Subtilus от вегетативной стадии к спорообразованию показали, что этот процесс во многом определяется изменением матричной активности РНК-полимеразы вегетативных клеток на раннем этапе споруляции. Показано, что во время вегетативного роста 85% иРНК транскрибируется с тяжелой нити ДНК и лишь 15% – с легкой нити. Популяция иРНК спорулирующих летокпредставлена иРНК, транскрибирующейся как с легкой, так и с тяжелой нити ДНК, причем с преимущественным синтезом иРНК на матрице легкой нити ДНК. На основании этих данных было сделано предположение, что споруляционные гены имеют специфичные промоторные участки на легкой нити ДНК /30/.
Показано, что бактериофаг b3 может инфицировать и реплицироваться в клетках Bac. subtilus, находящихся в логарифмической фазе роста, но не в спорулирующих клетках. В дальнейшем было обнаружено, что только РНК-полимеразы из клеток в логарифмической фазе обладает матричной специфичностью в отношении ДНКфага, тогда как РНК-полимераза спорулирующих клеток неспособна использовать ДНК данного фага как матрицу, но обладает повышенной активностью на синтетической матрице поли-dАТ. Вместе с тем подобного изменения матричной активности у мутантов с нарушенным спорообразованием в стационарной фазе роста не происходило. В связи с этим считают, что при переходе от вегетативного роста к спорообразованию происходит модификация специфичности РНК-полимеразы, следствием чего является прекращение считывания вегетативных членов и «включение» экспрессии споровых генов /31/.
Первоначально считали, что изменение специфичности РНК-полимеразы связано с модификацией b-субъединицы фермента, причем появление модифицированной субъединицы происходит за счет ее протеолитического расщепления щелочной протеиназы. Эти выводы были сделаны на основании того, что РНК-полимераза, выделенная из вегетативных и спорулирующих клеток, имела b-субъединицы с разным молекулярным весом. Таким образом, изменение матричной специфичности РНК-полимеразы не связано с модификацией минимального фермента, а, по мнению некоторых авторов, связано с утратой или инактивацией s-фактора /32/.
Таким образом, природа изменений РНК-полимеразы при переходе от вегетативных клеток к споруляции и роль этих изменений в ее матричной активности для включения процесса спорообразования окончательно не установлены. Вместе с тем данное направление исследования во многом определит выяснение природы перехода клеток к споруляции.
Регуляция спорообразования на уровне трансляции. Известно, что процесс спорообразования начинает осуществляться в стационарной стадии развития Bacillius, на которой происходят многочисленные изменения в аппарате трансляции клетки. В связи с этим определение изменений в данном аппарате, которые связаны именно со споруляционным процессом, затруднено. Это объясняется следующими причинами. Для обнаружения изменений в аппарате трансляции необходимо выделение, изучение и сравнение внутриклеточных компонентов, участвующих в трансляции, из вегетативно растущих клеток, клеток в стационарной фазе и спор. Однако в связи с тем, что количество нуклеаз и протеинах к моменту начала процесса спорообразования значительно возрастает, компоненты трансляционного аппарата претерпевают неспецифические модификации в момент своего выделения /33/.
До настоящего времени окончательно не установлено существование стабильных иРНК, специфичных для спорообразования. Однако имеются данные, указывающие на наличие стабильных споруляционных иРНК. Добавление меченого актиномицина D к культуре Bac. subtilus через определенные промежутки времени от начала процесса спорообразования позволило обнаружить различные классы спороспецифичных иРНК /32/.
Результаты изучения ДНК-РНК-гибридизации также указали на существование стабильной фракции иРНК в прикрепленных к мембране полирибосомах спорулирующих клеток, обработанных актиномицином D. Рядом исследователей показано, что при переходе к споруляции рибосомы вегетативных клеток претерпевают изменения. В результате использования белок-синтезирующей системы in vitro обнаружено, что у Bac. subtilus рибосомы становятся неспособными транслировать вегетативные иРНК при переходе клеток к споруляции. Таким образом, на основании данных, указывающих на наличие стабильной фракции иРНК, изменений в структуре рибосом, конформационных изменений в клеточной мембране, можно предполагать возможность функциональных изменений механизма, который определяет специфичную для процесса спорообразования трансляцию /31/.
Посттрансляционная регуляция спорообразования. Регуляция спорообразования на посттрансляционном уровне в настоящее время изучена недостаточно, однако считается, что этот вид регуляции, связанный с модификацией белков клетки, является необходимым для завершения споруляционного процесса. Обнаружен специфичный состав белков для каждой стадии спорообразования. Считается, модификация РНК-полимеразы, субъединиц рибосом и мембранных компонентов клетки происходит в результате посттрансляционных изменений и связана с фосфорилированием, аденилированием уже существующих белков, а также с образованием пептидных антибиотиков /27/.
Роль щелочной протеиназы в процессе спорообразования. В процессе споруляции Bac. subtilus образуются тир протеолитических активных фермента: сериновая протеиназа (субтилизин), работающая при щелочных значениях рН, в связи с чем в литературе обычно приводится название щелочная протеиназа; металлсодержащая протеиназа (нейтральная), рН оптимум которой находится в нейтральной зоне рН, и эстераза – фермент, обладающий невысокой протеолитической активностью. Различные представители рода Bacillus обладают специфической способностью к образованию определенного типа внеклеточной протеиназы. Так, Bac. megaterium и Bac. cereus синтезируют только нейтральную протеиназу, в то время как Bac. licheniformis, Bac. natto и Bac. subtilis образуют как нейтральную, так и щелочную протеиназы. Следует отметить, что Bac. subtilis по способности образования протеиназы различного типа можно подразделить на четыре группы; к первой относятся штаммы, синтезирующие только щелочную протеиназу, вторая группа образует только нейтральную протеиназу, представители третьей группы выделяют в среду как щелочную, так и нейтральную протеиназы, щелочная протеиназа у представителей четвертой группы появляется в среде только после того, как исчезнет нейтральная протеиназа /34/.
Установлено, что нейтральная протеиназа не участвует в процессе спорообразования, тогда как щелочная протеиназа необходима для включения данного процесса. В частности, выделен температурочувствительный по щелочной протеиназе мутант Bac. subtilis ts5. В условиях инкубации при 470С, т.е. температуре, при которой проявляется дефект температурочувствительного субтилизина, мутант не спорулировал. Восстановление активности щелочной протеиназы при 300 сопровождалось споруляцией вегетативно растущих клеток. Момент нарушения процесса спорообразования у данного ts5 мутанта совпадает с 0 стадией споруляции /35/.
В то же время описана большая группа плейотропно негативных мутантов, которые имели блок на 0 стадии спорообразования и характеризовались отсутствием способности к образованию щелочной протеиназы. Таким образом, из представленных данных можно судить о наличии корреляции между образованием щелочной протеиназы и спорообразованием. В последующих работах для определения участия протеиназ в процессе споруляции использовали два подхода. Первый подход состоял в создании условий, при которых ингибируется спорообразование. Обнаружено, что клетки, голодающие по тимидину, не спорулируют и не образуют такие экзоферменты, как протеиназы, рибонуклеазы, щелочную фосфатазу, при это не затрагивается только образование a-амилазы. На основании этих данных авторы предложили, что указанные ферменты прямо или косвенно связаны со спорообразованием /36/.
Вторым подходом для определения участия именно щелочной протеиназы в споруляционном процессе послужило использование специфичного ингибитора щелочной протеиназы – фенилметилсульфонилфторида (PMSF). Добавляя ингибитор через различные промежутки времени от начала процесса образования спор, обнаружили, что в осуществлении данного процесса имеется чувствительный период, продолжающийся 2–3 часа. Добавление ингибитора в этот период приводит к нарушению спорообразования, при этом не образуется щелочная фосфатаза, рибонуклеаза и термоустойчивые споры. Уровень синтеза нейтральной протеиназы в отсутствие ингибитора постоянно возрастает к началу спорообразования, затем либо незначительно снижается, либо достигает плато. Добавление PMSF приводит к возрастанию уровня накопления нейтральной протеиназы, при этом спорообразование отсутствует. На основании этих данных авторы сделали вывод о том, что нейтральные протеиназы по мере своего накопления разрушаются щелочной протеиназой и не участвуют в спорообразовании /26/.
Подтверждением того, что нейтральная протеиназа не принимает участия в процессе споруляции, послужили нормально спорулирующие мутанты Bac. subtilis, не обоазующие металлсодержащей протеиназы. Мутанты же, дефектные по образованию щелочной протеиназы, как правило – аспорогенны. Добавление ингибитора щелочной протеиназы после чувствительного периода не нарушает ни образования щелочной фосфатазы, ни появления светопреломляющих спор. Следовательно, щелочная протеиназа необходима для осуществления первых этапов спорообразования, в частности 0–1. Несмотря на то, что фермент продолжает накапливаться в течение всего периода спорообразования, для дальнейшего развития споры, по мнению авторов, щелочная протеиназа не явялется необходимой /27/.
Вместе с тем в ряде работ была показана важная роль щелочной протеиназы в модификации ферментов на поздних этапах спорообразования. Например, показано, что протеиназа может превращать фермент вегетативных клеток фруктозо-6-фосфатальдолазу в белок, специфичны для зрелых спор Bac. subtilis. Не исключено также возможное участие щелочной протеиназы в прорастании спор, при этом предполагают, что щелочные протеиназы, гидролизуюя белковую оболочку спор, включают их прорастание.
Все приведенные выше работы касаются роли внеклеточной щелочной протеиназы в спорообразовании.
Вместе с тем установлено, что к моменту начала процесса спорообразования Bac. subtilis образуется также и внутриклеточная щелочная протеиназа, уровень активности которой составляет 10% от соответствующего уровня внеклеточного фермента. Другие виды Bacillus, например Bac.megaterium, синтезируют, в отличие от Bac. Subtilis, толькл внутриклеточную щелочную протеиназу. В некоторых химических свойствах двух протеиназ существуют различия. В частности, указывается на наличие различной потребности в кофакторах и субстратной специфичности. Было также обнаружено, что двум щелочным протеиназам свойственна различная электрофоретическая подвижность. Внутриклеточная протеиназа обладает более узкой специфичностью в отношении эфиров и абсолютной потребностью в кальции. До последнего времени неясным остается вопрос о роли внутриклеточной протеиназы в спорообразовании Bac. Subtilis, хотя и описаны мутанты, у которых наряду с отсутствием активности внутриклеточной протеиназы не осуществляется процесс споруляции /34/.
Таким образом, процесс спорообразования регулируется на различных уровнях и образующиеся на разных этапах дифференцировки спор биологически активные соединения, в частности представляющие практический интерес, могут участвовать в регуляции этого процесса. В связи с эти принципы регуляции спорообразования представляют интерес. Однако в настоящее время еще мало известно о характере регуляции процесса спорообразования и регуляции синтеза соединений, специфичных для споруляции.
Похожие работы

... продукции, а значит, дает значительный экономический эффект. 2. Объекты и методы исследования Целью дипломной работы являлось изучение свойств бактериальной суспензии, с последующим применением в подготовительных процессах переработки мехового сырья. Для выполнения эксперимента был составлен сетевой график, представленный на рисунке 1. Применение ферментов в кожевенной и ...


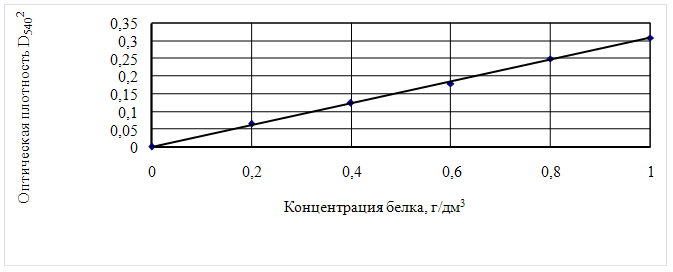
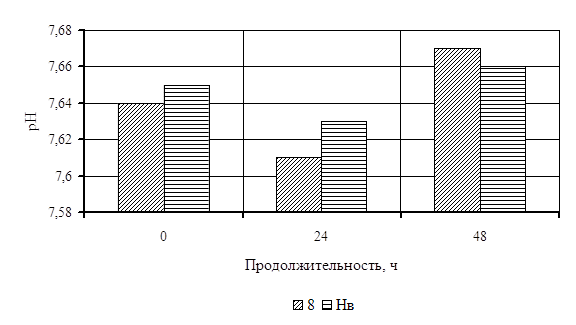
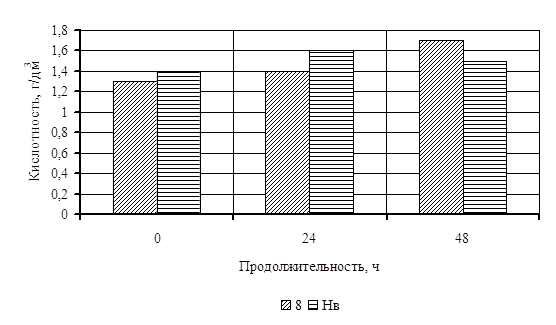
... Охрана окружающей среды Заключение Рисунок 2 – Сетевой график дипломной работы 2.1 Объекты исследования Объектом исследования в дипломной работе являлись микроорганизмы, выделенные из различных природных жиров: нерпичьего (Н), нерпичьего, выращенного на среде с шёрстным жиром (Нв), шерстного (В) и микроорганизмы, выделенные из ...
0 комментариев