Молекулярные структуры, ответственные за транспорт калия в МХ
АТФ-ингибируемые калий-транспортирующие каналы
АТФ-чувствительный калиевый канал внутренней мембраны МХ
Модуляторы митоКАТФ канала
Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ
Феномен прерывистой гипобарической тренировки
Выделение МХ печени крысы
Подготовка белка с м.м. 55 кДа: выделение и очистка
Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода
Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А
Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
Навигация
Молекулярные структуры, ответственные за транспорт калия в МХ
Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
118823
знака
4
таблицы
24
изображения
1.1.3 Молекулярные структуры, ответственные за транспорт калия в МХ
В настоящее время в литературе в качестве транспортных систем, осуществляющих электрогенный транспорт калия в МХ, рассматриваются следующие структуры: 1) АТФ/АДФ-антипортер или аденинуклеотидтранслоказа (АНТ) [Panov et al., 1980; Le Quoc et al., 1988] и 2) специфические белки, которые образуют во внутренней мембране МХ канал для электрогенного входа ионов калия [Миронова и др., 1981; Diwan et al., 1988; Paucek et a., 1992]. В первом случае предполагается, что АНТ, помимо основных функций, может работать как система электрофоретического транспорта ионов калия в МХ при действии на МХ субмикромолярных концентраций ионов кальция, и как неспецифическая пора для низкомолекулярных соединений при повреждении МХ высокими концентрациями Ca2+ [Jung and Brierley, 1981; 1982; Halestrap et al., 1986].
Что касается специфических белков, следует отметить, что в лаборатории Мироновой в 1981 г. методом водно-этанольной экстракции из МХ сердца быка был выделен и очищен белок с м.м. 55 кДа, который при реконструкции в бислойные липидные мембраны (БЛМ) образовывал К+-селективные каналы проводимости [Миронова и др., 1981], которые, как было показано позднее, ингибируются физиологической концентрацией АТФ и глибенкламидом, и относятся к семейству АТФ-чувствительных калиевых каналов [Миронова и др., 1996 (I); Mironova et al., 1999]. Структура митохондриального КАТФ канала не определена [Paucek et al., 1992].
Диван с соавторами, используя детергент Тритон Х-100, из МХ мембраны печени крыс выделили белок с м.м. 53 кДа [Diwan et al., 1988]. Реконструировав его в липосомы, авторы показали, что этот белок также обладает свойствами, характерными для К+ унипортера.
Известно, что в МХ помимо системы электрогеннного входа К+ существует система электронейтрального выхода К+ в обмен на Н+ [Garlid and Paucek, 2003]. При этом внешняя МХ мембрана не препятствует дальнейшему обмену небольшими ионами с цитоплазмой.
Система электрогенного входа калия и электронейтрального К+/Н+-обменника образуют К+ цикл МХ (Рис.1).
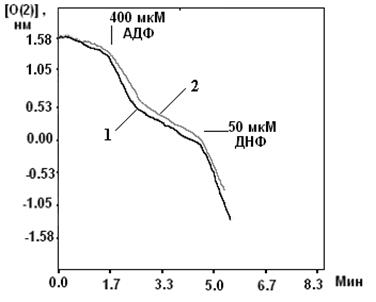
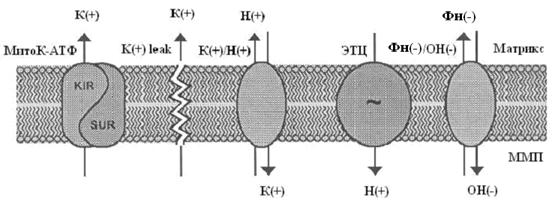
Рис. 1. МХ калиевый цикл по Garlid and Paucek, 2003. ЭТЦ – электрон-транспортная цепь; ММП – межмембранное пространство; Фн – неорганический фосфат.
Электрогенный выброс протонов электрон-транспортной системой генерирует мембранный потенциал, который, в свою очередь, стимулирует диффузию К+ в матрикс МХ («утечка калия» или К+ leak) и вход иона, опосредованный неким специфическим калиевым каналом. Такой обмен Н+ на К+ подщелачивает матрикс, вызывая вход фосфата по электронейтральному Фн-Н+ симпортеру (Фн – неорганический фосфат) (Рис.1).
Вход К+ сопровождается накоплением осмотически облигатной воды, что проявляется в набухании МХ. Излишнее их набухание может угрожать целостности этих органелл, следовательно, избыток К+ необходимо удалять. Эту функцию выполняет белок с м.м. 82 кДа - К+/Н+-антипортер [Brierley G., 1976; Garlid K., et al., 1980; 1988]. Следует отметить, что вход К+ путем диффузии слишком мал, чтобы существенно влиять на изменение объема матрикса МХ. Интенсивный вход калия в МХ обеспечивается специфическими белками, осуществляющими электрофоретический вход К+ в МХ [Mironova G., 1981; Diwan J., et al., 1988; Paucek et al., 1992].
1.1.3 Физиологическое значение транспорта калия в МХПредполагается, что основное физиологическое значение систем транспорта К+ в МХ связано с регуляцией объема МХ матрикса. Объем МХ определяется калиевыми токами через внутреннюю мембрану. Когда вход и выход К+ находятся в равновесии, калиевые потоки определяются электронейтральным током анионов и осмотически облигатной воды [Garlid, 1988]. Так как концентрация ионов калия в матриксе и в цитоплазме практически одинаковая, транспорт калия мало влияет на матриксную концентрацию калия, но может иметь большое влияние на объем МХ. Небольшое нескомпенсированное увеличение входа К+ в МХ может удваивать их объем в течение 1-2 минут [Garlid, 1979]. В свою очередь, увеличение матриксного объема стимулирует активность дыхательной цепи, что было показано на МХ сердца и печени [Nichols and Lindberg, 1972; Halestrap, 1989]. Окисление жирных кислот также чувствительно к изменению объема матрикса [Halestrap, 1987].
Увеличение в МХ концентрации К+ и последующее изменение их объема имеет значение и в регуляции митохондриальных процессов у зимоспящих животных [Fedotcheva et al., 1985; Бакеева и Брустовецкий, 1993], а также при адаптации животных к холоду [Nedergraard and Cannon, 1987]. В ряде работ установлено, что in vivo, гормон, участвующий в регуляции теплопродукции (тироксин), активирует системы электрогенного транспорта К+, вызывая низкоамплитудное набухание МХ [Halestrap, 1987; Shears an Brouk, 1980]. В нашей лаборатории, было показано, что активность системы электрогенного транспорта ионов калия прямо коррелирует с интенсивностью термогенеза [Федотчева и др., 1984; Миронова и др., 1986], что непосредственно связано с увеличением содержания ионов калия в МХ бурой жировой ткани и печени [Скарга, 1994]. Активацию транспорта калия при выходе животного из спячки, связывают с активацией футильного цикла К+ в МХ, приводящей к увеличению теплопродукции, до начала синтеза АТФ в МХ [Миронова и др., 1986].
Обнаружено также, что митоКАТФ канал играет существенную роль в нормальной физиологии миокарда, регулируя объем митохондрий и продукцию активных форм кислорода (АФК) [Garlid et al., 2003a].
В последнее время интерес ученых к митоКАТФ каналу вызван обнаружением его роли в защите сердца от инфаркта. Исследования были стимулированы открытием феномена кардиопротекторного действия прерывистой гипоксии, приводящей к активации митоКАТФ [Murry et al., 1986]. Установлено, что фармакологические активаторы митоКАТФ канала предохраняют сердце от ишемических повреждений [Garlid et al., 1997].
0 комментариев