Молекулярные структуры, ответственные за транспорт калия в МХ
АТФ-ингибируемые калий-транспортирующие каналы
АТФ-чувствительный калиевый канал внутренней мембраны МХ
Модуляторы митоКАТФ канала
Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ
Феномен прерывистой гипобарической тренировки
Выделение МХ печени крысы
Подготовка белка с м.м. 55 кДа: выделение и очистка
Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода
Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А
Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
Навигация
Модуляторы митоКАТФ канала
Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
118823
знака
4
таблицы
24
изображения
2.1 Модуляторы митоКАТФ канала
2.1.1 Метаболические модуляторы митоКАТФ канала
В связи с выраженным кардиопротекторным действием активаторов митоКАТФ в настоящее время во многих лабораториях ведется поиск новых синтетических активаторов канала, которые могут быть потенциальными кардиопротекторами [Ockaili et al., 2002; Kopustinskene et al., 2001]. В нашей лаборатории помимо синтетического донора электронов ДЕБ, который селективно активировал митоКАТФ [Mironova et al., 1999; Mironova et al., 2004], был обнаружен ряд природных, то есть метаболических активаторов этого канала.
Дифосфонуклеотиды
Показано, что дифосфонуклеотиды (АДФ и ГДФ) активируют реконструированную в БЛМ канальную субъединицу (Рис. 2) [Grigoriev et al., 1999; Mironova et al., 1999]. Данные по активирующему действию ГДФ в дальнейшем были подтверждены и другими исследователями [Bednarczyk et al., 2005]. Недавно в нашей лаборатории было показано, что в ряду дифосфонуклеотидов наиболее эффективным является уридиндифосфат (УДФ) [Mironova et al., 2004].
В МХ УДФ (~20 мкМ) активирует митоКАТФ, и эта активация снимается глибенкламидом и 5-ГД (Рис. 4) [Mironova et al., 2004]. Концентрации УДФ, неоходимые для активации реконструированного в БЛМ митоKIR и целого канала в нативных МХ, одинаковы. Следовательно, участок связывания УДФ локализуется на канальной субъединице. Для активации цитоКАТФ нужны значительно большие концентрации этого препарата [Alekseev et al., 1998].
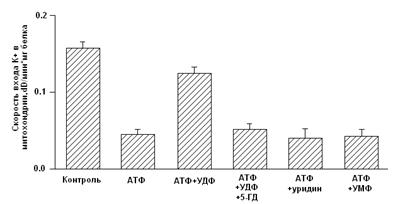
Рисунок 4. Активация УДФ целого митоКАТФ канала в нативных МХ [Mironova et al., 2004]
Ранее на моделях тотальной и регионарной ишемии изолированных перфузируемых сердец крыс было показано, что уридин и УМФ проявляют антиаритмическую активность в отношении реперфузионных аритмий, предупреждают развитие миокардиального станинга при постишемической реперфузии, препятствуя депрессии сократительной функции миокарда и предотвращая развитие контрактуры [Eliseev et al., 1989].
В представленной работе была предпринята попытка определить, обладают ли уридиновые нуклеотиды кардиопротекторным действием и связано ли их влияние с активацией цито- и митоКАТФ каналов на модели искусственного острого инфаркта миокарда.
2.2 Функциональная роль митоКАТФ
Являясь одним из компонентов калиевого цикла МХ, митоКАТФ участвует в поддержании объема МХ матрикса и мембранного потенциала [Garlid, 1998]. В лаборатории проф. Мироновой показано, что активация митоКАТФ играет важную роль при выходе животных из состояния зимней спячки [Fedotcheva et al., 1985; Миронова и др., 1986]. МитоКАТФ участвует также в ингибировании процесса апоптоза клетки, блокируя высвобождение цитохрома С, предотвращая снижение мембранного потенциала и чрезмерное увеличение концентрации Ca2+, сохраняя, таким образом, функции МХ, что было показано на b-клетках поджелудочной железы [Akao et al., 2001], а затем и на церебральных нейронах [Teshima et al., 2003]. Кроме того, митоКАТФ является посредником в процессе регуляции транскрипции некоторых генов. Его активация вызывает небольшое увеличение продукции активных форм кислорода (АФК) [Chandel et al., 1998].
При патологии активация митоКАТФ частично востанавливает мембранный потенциал, стимулируя дальнейший перенос протонов, ингибируя накопление внутримитохондриального кальция и предотвращая распад энергетических фосфатов путем создания оптимального для синтеза АТФ электрохимического градиента [Szewczyk, 1996; Xu et al., 2001]. Активация митоКАТФ способствует разобщению электронного транспорта, ускоряя, таким образом, дыхание, что приводит к окислению проксимальных переносчиков электронов дыхательной цепи [Liu et al., 1998]. Согласно данным Гарлида, активация канала приводит к набуханию МХ, что способствует поддержанию архитектуры внутренней мембраны и сохранению активности дыхательной цепи. Это позволяет поддерживать уровень клеточного АТФ и, таким образом, способствует восстановлению функций миокарда во время реперфузии [Garlid, 2000]. Как было показано ранее, набухание матрикса увеличивает скорость окислительного метаболизма, активирует как окисление жирных кислот, так и дыхание и синтез АТФ [Halestrap, 1989].
2.2.1 Активация митоКАТФ в развитии устойчивости организма к гипоксииВ 1983 г. был открыт феномен прерывистой гипоксии, важную роль в котором играет активация митоКАТФ [Murry et al., 1983; Liu et al., 1998].
Феномен прерывистой гипоксии был назван «preconditioning» или «ишемической толерантностью» [Murry et al., 1986; Kitigawa, 1990]. Позднее он получил название «феномен ишемической адаптации» (ФИА) [Yellon et al., 1998; Петрищев и др., 2001], и в настоящее время признан наиболее эффективным способом уменьшения размера инфаркта миокарда [Kloner et al., 1998]. Справедливости ради надо отметить, что адаптация, создаваемая кратковременной гипоксией, была обнаружена ранее [Poupa et al.,1966; Meerson et al., 1973].
Клеточные механизмы ФИА в настоящее время широко изучаются. Считается, что защитный эффект является трехступенчатым и включает в себя последовательные триггерные и конечные эффекторные механизмы, причем наиболее вероятным эффектором является митоКАТФ [Garlid et al., 1997; Yellon et al., 1998; Grover and Garlid., 2000; Петрищев и др., 2001].
ФИА развивается в виде двух волн: первая волна проявляется в течение нескольких часов, затем исчезает и снова появляется в виде второй волны [Yellon et al., 1998], которая длится в течение недели, а иногда и месяца [Carroll et al., 2000]. Установлено, что во второй стадии адаптации к гипоксии также участвует митоКАТФ [Wang et al., 2001]. В развитии толерантности к ишемии мозга [Lacza et al., 2003], и в дистанционной ишемической адаптации сердца к гипоксии при создании коротких эпизодов окклюзии почечной артерии также задействован митоКАТФ [Pell et al., 1998].
Установлено, что специфические модуляторы митоКАТФ (см. таблицу 1) влияют на ФИА. Так активатор митоКАТФ, диазоксид, и неселективный активатор КАТФ каналов, кромакалим, имитируют «preconditioning», увеличивая время сокращения миокарда и способствовуя восстановлению функций изолированных сердец крыс, подвергавшихся общей ишемии миокарда с последующей реперфузией. Кi диазоксида для митоКАТФ (0.8 мкМ в МХ сердца быка) в 1000 раз превышала Кi для цитоКАТФ [Gross et al., 1992]. Кромакалим проявлял аналогичный эффект в тех же концентрациях. Глибенкламид и 5-ГД блокировали действие диазоксида и кромакалима, следовательно, эффект активаторов на ФИА опосредован через КАТФ каналы [Auchampach, 1992; Garlid, 1997; Tsai et al., 1998; Vanden Hoek, 2000]. Активирующе действие диазоксида на митоКАТФ показано также на изолированных МХ сердца крысы, что доказывает роль МХ канала в кардиопротекции [Gross et al., 1992]. Полученные данные подтверждают также результаты исследований, по изучению флуоресценции флавопротеинов на изолированных кардиомиоцитах желудочков сердца кроликов [Liu et al., 1998; Sato et al., 1998]. Однако поскольку было обнаружено, что диазоксид снижает мембранный потенциал на МХ, независимо от активации митоКАТФ канала, его кардиопротекторный эффект вызывает сомнение.
Показано, что потенциальный медиатор сигнального пути, ответственного за формирование ишемической адаптации, протеинкиназа С, [Sato et al., 1998; Liu et al., 1994; Sasaki et al., 2000] модулирует активность митоКАТФ. 5-ГД полностью блокирует этот кардиопротекторный эффект, подтверждая участие митоКАТФ в защите миокарда от ишемических повреждений. Следует отметить, что сами по себе глибенкламид и 5-ГД не оказывают никакого влияния [Gross and Fryer, 1999]. Неспособность специфического ингибитора цитоКАТФ, HMR-1883, ингибировать анти-ишемический эффект активаторов митоКАТФ [Fryer et al., 2000], также говорит в пользу участия в защитном механизме МХ канала.
Следует отметить, что неселективные активаторы КАТФ каналов, такие как пинацидил, оказывая кардиопротекторное действие, вызывали побочный проаритмический эффект и приводили к существенному понижению давления у подопытных животных [Grover, 1997]. Подобные нежелательные последствия способны свести на нет кардиопротекторное действие данных препаратов. Использование селективных активаторов МХ или цитоплазматического КАТФ каналов позволило бы избежать таких побочных явлений. Можно ожидать, что селективные активаторы цитоКАТФ, такие как HMR-1883, оказывая антиаритмическое действие, не будут негативно влиять на ишемическую адаптацию, вызванную активацией митоКАТФ. Так, показано, что HMR-1883 является потенциальным антифибрилляторным агентом при острой ишемии на модели собаки [Billman et al., 1998]. И наоборот, сайт-специфичные активаторы митоКАТФ не будут оказывать проаритмического действия [Gross and Fryer, 1999].
0 комментариев